- Trichoplax
-
Trichoplax adhaerens 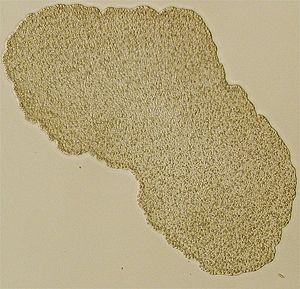
Lichtmikroskopisches Bild von Trichoplax (Individuum ca. 0,5 mm im Durchmesser)
Systematik Reich: Tiere (Animalia) Unterreich: Vielzellige Tiere (Metazoa) Abteilung: Gewebelose (Parazoa) Stamm: Placozoa Gattung: Trichoplax Art: Trichoplax adhaerens Wissenschaftlicher Name Trichoplax adhaerens F. E. Schulze 1883 Trichoplax adhaerens ist die einzige bekannte Art der Placozoa und wurde 1883 von dem deutschen Zoologen Franz Eilhard Schulze in einem Meerwasser-Aquarium des Zoologischen Instituts in Graz entdeckt. Der von ihm verliehene Gattungsname des Tiers leitet sich aus dem altgriechischen θρίξ thrix „Haar“ und πλάξ plax „Platte“ ab, das lateinische Artepithet adhaerens bedeutet „haftend“, so dass sich der Artenname etwa mit „anhaftende haarige Platte“ übersetzen lässt.
Die Placozoa sind die strukturell einfachsten aller vielzelligen Tiere (Metazoa) und bilden mit nur einer einzigen Art, Trichoplax adhaerens, einen eigenen Tierstamm. Ein deutscher Name für die Gruppe existiert noch nicht; der wissenschaftliche Name bedeutet wörtlich übersetzt „flache Tiere“, vielfach wird im Deutschen der Name „Scheibentiere“ verwendet.[1]
Lange Zeit hielt man die bisher nicht in ihrem natürlichen Lebensraum beobachteten Tiere für Jugendstadien von Nesseltieren (Cnidaria): Länger als ein halbes Jahrhundert wurden sie als Planula-Larven der Hydrozoen-Art Eleutheria krohni gedeutet. Erst Arbeiten aus den 1970er Jahren, unter anderem durch den Tübinger Protozoologen Karl Gottlieb Grell, klärten die frühen Phasen der Embryonalentwicklung der Tiere auf und trugen maßgeblich zur Errichtung eines eigenen Tierstamms für sie bei. Formell wird die Art der Familie Trichoplacidae zugeordnet.
Inhaltsverzeichnis
Aufbau
Placozoa haben, wie ihr Name andeutet, in der Regel einen stark abgeflachten, scheibenförmigen Körper. Er misst im Durchmesser oft weniger als einen halben, selten aber mehr als zwei bis drei Millimeter; die Scheibendicke beträgt meistens nur etwa 25 Mikrometer. Mit bloßem Auge sind die gräulich gefärbten, im Gegenlicht transparenten Tiere meist gerade noch zu erkennen. Oberflächlich sehen sie wie große Amöben aus und ändern wie diese beständig ihre äußere Form. Selten treten daneben kugelförmige Lebensstadien auf, die möglicherweise dem passiven Transport in neue Lebensräume dienen.
Wie die Zuordnung zu den Gewebelosen schon andeutet, fehlen Trichoplax adhaerens Gewebe und Organe; es gibt auch keine ausgeprägte Körpersymmetrie, so dass sich weder vorne oder hinten noch links oder rechts unterscheiden lassen.
Epitheloid
Sowohl strukturell als auch funktionell lassen sich bei Trichoplax adhaerens eine Rücken- oder Dorsalseite und eine Bauch- oder Ventralseite unterscheiden. Beide bestehen aus einer einlagigen, nach außen von Schleim bedeckten Zellschicht und erinnern vor allem wegen der zwischen den Zellen bestehenden Verbindungen, der Gürteldesmosomen, an Epithelgewebe. Die Zelllagen der Placozoa besitzen aber im Gegensatz zu einem echten Epithel keine Basallamina; darunter versteht man eine aus extrazellulärem Material bestehende, dem Epithel von innen unterliegende dünne Schicht, die es versteift und vom Innenraum des Körpers abtrennt. Das Fehlen dieser ansonsten bei allen Tieren außer den Schwämmen zu findenden Struktur lässt sich funktionell begründen: Eine steife Trennschicht würde die amöboiden Formänderungen von Trichoplax adhaerens unmöglich machen. Statt von einem Epithel spricht man daher bei den Placozoa von einem Epitheloid.
Ein ausgewachsenes Individuum besteht aus bis zu tausend Zellen, die vier verschiedenen Zelltypen zugeordnet werden können. Die einfach begeißelten Zellen des rückseitigen Epitheloids sind abgeflacht und enthalten kleine Lipidkörperchen. Die Zellen der Bauchseite sind zwar ebenfalls nur einfach begeißelt, haben aber eine langgestreckte säulige Form mit kleinem Querschnitt an der Oberfläche, so dass die Geißeln bauchseitig sehr eng beieinander stehen und eine bewimperte „Kriechsohle“ bilden. Dazwischen befinden sich unbewimperte Drüsenzellen, die vermutlich in der Lage sind, Verdauungsenzyme zu synthetisieren.
Fasersynzytium
Zwischen den beiden Zellschichten befindet sich ein flüssigkeitsgefüllter Innenraum, der mit Ausnahme der unmittelbaren Kontaktzone von Vorder- und Rückseite von einem sternförmigen Fasersynzytium durchsetzt ist: Darunter versteht man ein faseriges Netzwerk, das effektiv aus einer einzigen Zelle besteht, aber zahlreiche Zellkerne enthält, die zwar durch interne Querwände (Septa), nicht aber durch echte Zellmembranen voneinander getrennt sind. Ähnliche Strukturen findet man auch bei Schwämmen (Porifera) und vielen Pilzen (Fungi).
Auf beiden Seiten der Septa befinden sich flüssigkeitsgefüllte Kapseln, durch welche die angeführten Trennstrukturen Synapsen ähneln, also Nervenzellverbindungen, die sich in voll ausgeprägter Form nur bei Gewebetieren (Eumetazoa) finden. Auffällige Ansammlungen von Kalzium-Ionen, die eine Funktion bei der Erregungsweiterleitung haben könnten, weisen ebenfalls auf eine mögliche Rolle als Protosynapsen hin. Zu dieser Ansicht passt, dass sich fluoreszierende Antikörper gegen Nesseltier-Neurotransmitter, also gerade die in Synapsen übertragenen Signalstoffe, konzentriert in bestimmten Zellen von Trichoplax adhaerens binden und damit auf die Existenz vergleichbarer Substanzen bei den Placozoa hindeuten. Das Fasersynzytium enthält daneben Aktin- und vermutlich auch Myosin-Moleküle, die bei Gewebetieren in den Muskelzellen vorkommen. Bei den Placozoa sorgen sie dafür, dass sich die einzelnen Fasern entspannen oder zusammenziehen können, und wirken so an der Formbildung der Tiere mit.
Auf diese Weise übernimmt das Fasersynzytium funktionell sowohl die Aufgaben von Nerven- als auch von Muskelgewebe. Außerdem findet hier zumindest ein Teil der Verdauung statt. Andererseits existiert keine gallertige extrazelluläre Matrix wie man sie bei Nesseltieren oder Rippenquallen als Mesoglea findet.
Anders als bei den Gewebetieren wurden pluripotente Zellen, also solche, die sich in andere Zelltypen umwandeln können, noch nicht eindeutig nachgewiesen. Rückseitige und bauchseitige Epithelzellen gehen nach traditioneller Ansicht immer nur aus ihresgleichen hervor.
Genetik
Alle Zellkerne der Placozoa-Zellen enthalten zwölf nur etwa zwei bis drei Mikrometer große Chromosomen. Drei Paare sind metazentrisch, die anderen acrozentrisch, das heißt, das Zentromer, die Ansatzstelle der Spindelfasern bei der Zellteilung, liegt entweder in der Mitte oder am äußeren Rand des jeweiligen Chromosoms. Die Zellen des Fasersynzytiums sind eventuell tetraploid, enthalten also einen vierfachen Chromosomensatz.
Insgesamt enthält ein einfacher Chromosomensatz von Trichoplax adhaerens weniger als 50 Millionen Basenpaare und bildet damit das kleinste tierische Genom; die Zahl der Basenpaare des Darmbakteriums Escherichia coli ist nur um einen Faktor 10 kleiner.
Die genetische Ausstattung von Trichoplax adhaerens ist noch nicht sehr gut erforscht; einige Gene wie Brachyury oder Tbx2/3, die homolog zu entsprechenden Basenpaarsequenzen bei Gewebetieren sind, konnten allerdings bereits nachgewiesen werden. Von besonderer Bedeutung ist Trox-2, ein Placozoa-Gen, das bei Nesseltieren unter dem Namen Cnox-2 und bei den zweiseitig-symmetrischen Bilateria als GSX bekannt ist. Als Homöobox- oder HOX-Gen spielt es in der Embryonalentwicklung bei Gewebetieren eine Rolle bei der Ausbildung und Körperdifferenzierung entlang der Symmetrieachse; bei den Nesseltieren legt es etwa die Lage von Mund- (oraler) und mundabgewandter (aboraler) Seite des Tieres fest. Placozoa besitzen allerdings wie angesprochen keine Symmetrieachsen. Daher ist es von besonderem Interesse, wo genau im Trichoplax-Körper das Gen abgelesen wird. Durch Antikörperstudien konnte nachgewiesen werden, dass das Genprodukt nur in der Übergangszone von Rücken- und Bauchseite vorkommt, vielleicht in einem bisher noch nicht charakterisierten fünften Zelltyp. Ob es sich bei diesen Zellen im Gegensatz zu traditionellen Vorstellungen um Stammzellen handelt, die eine Rolle bei der Zelldifferenzierung spielen, ist noch unklar. In jedem Fall gilt Trox-2 als möglicher Kandidat für ein Proto-HOX-Gen, aus dem durch Verdopplungen und Differenzierung die anderen Gene dieser wichtigen Gen-Familie entstanden sein könnten.
Mittels molekularbiologischer Methoden wurde zunächst erfolglos versucht, die verschiedenen Theorien zur Stellung der Placozoa im System der Vielzeller zu testen. Mit Standardmarkern wie 18S-rDNA/RNA konnte keine Klärung erzielt werden: Die Markersequenz war offenbar „verrauscht“, also durch zu viele Mutationen uninformativ geworden. Immerhin stützte dieses Negativergebnis den Verdacht, dass Trichoplax eine äußerst urtümliche Vielzellerlinie darstellen könnte, da für die Anhäufung der zahlreichen Mutationen ein sehr langer Zeitraum angenommen werden musste.
Die eigentlich interessierende Frage, ob die Placozoa älter sein könnten als die Schwämme, scheint neuerdings durch Aufklärung des mitochondrialen Genoms beantwortet zu sein. Das mitochondriale Genom weist Trichoplax als deutlich urtümlicher als die Schwämme aus, da es insgesamt eher an die Verhältnisse bei Einzellern erinnert (Dellaporta et al. 2006). Demnach wären die Placozoa also relativ kurz nach dem evolutionären Einzeller-Vielzeller-Übergang entstanden. Dieses Resultat bestätigt insbesondere eine auf der funktionsmorphologischen Interpretation des Placozoen-Bauplanes begründete Abstammungshypothese, und widerlegt gleichzeitig die bisherigen rein merkmalsmorphologischen Deutungen, in denen die Schwämme für älter gehalten wurden (zu den konkurrierenden morphologischen Theorien siehe den Abschnitt „Stammesgeschichte“).
Verbreitung und Lebensraum
Genaue Verbreitungsangaben existieren für die Placozoa nicht, die Tiere wurden aber unter anderem im Roten Meer, dem Mittelmeer, der Karibik, vor Hawaii, Guam, Samoa, Japan, Vietnam oder Papua-Neuguinea und am Großen Barriere-Riff vor der australischen Ostküste nachgewiesen. Regelmäßig wird Trichoplax adhaerens auch in Meerwasser-Aquarien „gefunden“, so etwa im südenglischen Plymouth oder im US-amerikanischen Bundesstaat Florida in Miami.
Die bisherigen Feldfunde stammen aus der küstennahen Gezeitenzone tropischer und subtropischer Meere, wo sich die Tiere auf Substraten wie Stämmen oder Wurzeln von Mangroven, Weichtier-Schalen, Bruchstücken von Steinkorallen oder einfach auf Felsbrocken finden. Eine Studie konnte jahreszeitliche Populationsschwankungen ausmachen, deren Ursachen allerdings bisher noch ungeklärt sind.
Ernährung und Symbionten
 Extrakorporale Nahrungsaufnahme von Trichoplax adhaerens
Extrakorporale Nahrungsaufnahme von Trichoplax adhaerensTrichoplax adhaerens ernährt sich von kleinen Algen, insbesondere Grünalgen (Chlorophyta) der Gattung Chlorella, Kryptomonaden (Cryptophyta) der Gattungen Cryptomonas oder Rhodomonas, Cyanobakterien (Cyanobacteria) wie Phormidium inundatum, aber auch von abgestorbenen Teilen anderer Organismen. Dazu bilden sich auf der Bauchseite um die Nahrungspartikel eine oder mehrere kleine Taschen aus, in die von den Drüsenzellen Verdauungsenzyme abgegeben werden; die Tiere entwickeln also gewissermaßen einen vorübergehenden „Außenmagen“. Die aufgeschlossenen Nährstoffe werden dann durch Pinocytose („Zell-Trinken“) von den bauchseitig gelegenen Geißelzellen aufgenommen.
Auch über das obere Epitheloid (also die „Rückenseite“ des Tieres) können Nahrungspartikel, z. B. ganze Einzeller, aufgenommen werden. Diese Ernährungsweise dürfte im Tierreich einmalig sein: Die in einer Schleimschicht gesammelten Nahrungspartikel werden von den Faserzellen durch Interzellularspalten (Zellzwischenräume) des Epitheloids hindurchgezogen und dann per Phagocytose („Zell-Essen“) verdaut. Ein solches „Einsammeln“ von Nahrungspartikeln durch ein intaktes Abschlussgewebe hindurch ist nur deshalb möglich, weil bei den Placozoa verschiedene „abdichtende“ Elemente (nämlich eine Basallamina unter dem Epitheloid und bestimmte Zell-Zell-Verbindungen) nicht vorhanden sind.
Nicht alle Bakterien, die sich im Inneren der Placozoa finden, werden als Nahrung verdaut: Im endoplasmatischen Reticulum, einem Zellorganell des Fasersynzytiums, befinden sich oft Bakterien, die mit Trichoplax adhaerens anscheinend in Symbiose leben.
Fortbewegung
Auf fester Oberfläche können sich Placozoa auf zwei verschiedene Arten bewegen: Einmal ermöglicht es ihnen ihre begeißelte Kriechsohle, langsam über den Untergrund hinweg zu gleiten, zum anderen können sie wie eine Amöbe ihre Lage durch Wandlung der Körperform verändern. Die Bewegungen sind dabei nicht zentral koordiniert, da keinerlei Muskel- oder Nervengewebe existiert. Es kann daher vorkommen, dass sich ein Individuum gleichzeitig in zwei verschiedene Richtungen bewegt und sich in Folge in zwei Teile spaltet.
Zwischen Körperform und Fortbewegungsgeschwindigkeit konnte eine enge Verbindung nachgewiesen werden, die zusätzlich abhängig vom Nahrungsangebot ist:
- Bei niedriger Nährstoffdichte schwankt die Querschnittsfläche geringfügig, aber unregelmäßig, die Fortbewegungsgeschwindigkeit liegt verhältnismäßig konstant bei etwa 15 Mikrometern pro Sekunde.
- Ist die Nährstoffdichte dagegen hoch, oszilliert der Querschnitt mit einer stabilen Periode von etwa 8 Minuten, wobei die größte Ausdehnung des Tiers bis zu doppelt so groß sein kann wie die kleinste. Die Fortbewegungsgeschwindigkeit, die durchweg unterhalb von 5 Mikrometern pro Sekunde liegt, schwankt mit derselben Periode. Eine große Geschwindigkeit entspricht dabei immer einer kleinen Querschnittsfläche und umgekehrt.
Die beiden Ausbreitungsweisen lassen sich, da der Übergang nicht flüssig, sondern abrupt vonstatten geht, sehr gut gegeneinander abgrenzen. Vereinfachend lässt sich Trichoplax adhaerens als nicht-lineares dynamisches System fern dem thermodynamischen Gleichgewicht modellieren.
Eine qualitative Erklärung für das Verhalten des Tiers ergibt sich wie folgt:
- Bei niedriger Nährstoffdichte behält Trichoplax eine konstante Geschwindigkeit bei, um ohne unnötigen Zeitverlust Nahrungsquellen ausfindig zu machen.
- Ist eine solche Quelle durch hohe Nährstoffdichte identifiziert, erhöht das betreffende Individuum in regelmäßigen Abständen seinen Querschnitt und vergrößert damit die Kontaktfläche mit dem Substrat. Auf diese Weise erweitert sich die Fläche, über die Nährstoffe aufgenommen werden können. Gleichzeitig verringert das Tier seine Geschwindigkeit, um das vorhandene Nahrungsangebot auch tatsächlich auszuschöpfen.
- Sobald dies annähernd geschehen ist, verkleinert Trichoplax seinen Querschnitt wieder, um sich weiterzubewegen. Weil Nahrungsquellen wie Algenteppiche oft eine größere Ausdehnung haben, ist es für ein betroffenes Tier sinnvoll, nach kurzer Zeit in der Bewegung innezuhalten, um sich erneut abzuflachen und Nährstoffe aufzunehmen. Daher bewegt sich Trichoplax in dieser Phase nur verhältnismäßig langsam voran.
Die konkrete Richtung, in die sich Trichoplax jeweils bewegt, ist zufallsbestimmt: Misst man, wie schnell sich ein einzelnes Tier von einem (willkürlich bestimmten) Ausgangspunkt wegbewegt, so findet man eine lineare Beziehung zwischen der vergangenen Zeit und der mittleren quadratischen Entfernung von Start- und aktuellem Aufenthaltsort. Ein solcher Zusammenhang charakterisiert auch die zufallsbestimmte Brownsche Bewegung, die somit als Modell für die Fortbewegung der Placozoa dienen kann.
Kleine Tiere sind auch in der Lage, aktiv mit Hilfe ihrer Geißeln zu schwimmen. Sobald sie ein mögliches Substrat berühren, kommt es zu einer Dorsoventralreaktion: Die Geißeln der Rückseite schlagen weiter, während die Geißeln der bauchseitigen Zellen ihren Schlagrhythmus einstellen. Gleichzeitig versucht die Bauchseite, Kontakt mit dem Untergrund herzustellen; kleine Zellaus- und -einstülpungen, die Mikrovilli, die sich an der Oberfläche der säuligen Zellen der Bauchseite befinden, tragen durch ihre adhäsive (klebende) Wirkung zur Anheftung an das Substrat bei.
Regeneration
Eine bemerkenswerte Eigenschaft der Placozoa ist, dass sie sich aus kleinsten Zellbeständen regenerieren können. Selbst wenn im Experiment große Teile des Organismus entfernt werden, entwickelt sich aus dem Rest wieder ein vollständiges Tier. Es ist darüber hinaus möglich, Trichoplax adhaerens durch ein Passiersieb zu streichen, so dass die Einzelzellen zwar nicht zerstört, aber weitgehend voneinander getrennt werden. Im Reagenzglas finden sie sich dann wieder zu kompletten Organismen zusammen. Wird diese Prozedur mit mehreren vorher angefärbten Tieren gleichzeitig vollzogen, geschieht das Gleiche. In diesem Fall kann es jedoch vorkommen, dass Zellen, die vorher zu einem bestimmten Tier gehörten, plötzlich als Teil eines anderen wieder auftauchen.
Fortpflanzung
Normalerweise pflanzen sich die Placozoa ungeschlechtlich fort. Das Tier schnürt sich dazu in der Mitte durch, so dass zwei etwa gleich große Tochtertiere entstehen, die nach der Fission allerdings noch für eine Weile lose verbunden bleiben. Seltener findet man Knospungsvorgänge: Dabei trennen sich von der Rückenseite kleine Zellkugeln ab, die alle bekannten Zelltypen in sich vereinigen und in der Folge zu einem eigenen Individuum heranwachsen.
Die geschlechtliche Fortpflanzung wird vermutlich durch eine zu große Populationsdichte ausgelöst. Sie wird dadurch eingeleitet, dass die Tiere Flüssigkeit aufnehmen, anschwellen und sich vom Substrat lösen, so dass sie frei im Wasser schweben. Die Zellen der Bauchseite bilden nun im geschützten Innenraum eine von einer speziellen Hülle, der Befruchtungsmembran, umgebene Eizelle aus, die durch das umgebende Synzytium mit Nährstoffen versorgt wird, so dass sich in ihrem Inneren energiereicher Dotter ansammeln kann. Sobald die Reifung der Eizelle abgeschlossen ist, degeneriert der Rest des Tieres, wodurch die Eizelle selbst freigesetzt wird. Kleine unbegeißelte Zellen, die sich gleichzeitig bilden, werden als Spermien interpretiert. Die Befruchtung selbst konnte bisher noch nicht beobachtet werden; die Existenz der Befruchtungsmembran wird aber gemeinhin als Beleg dafür angesehen, dass eine solche stattgefunden hat.
Die Eizelle beginnt, meist schon vor ihrer Freisetzung, mit Furchungsteilungen, bei denen sie sich vollständig in der Mitte durchschnürt. Auf diese Weise entsteht schließlich eine für Tiere charakteristische Zellkugel, die Blastula, die maximal 64 Zellen enthält. Die Entwicklung nach diesem 64-Zell-Stadium ist derzeit noch unbekannt.
Wegen der Möglichkeit, sich unbegrenzt durch ungeschlechtliche Fortpflanzung zu klonen, ist die Lebensspanne der Placozoa potentiell unendlich; in der Praxis werden einige Entwicklungslinien, die auf ein einziges Tier zurückgehen, seit mittlerweile 20 Jahren ohne das Auftreten sexueller Prozesse in Kultur gehalten.
Rolle als Modellorganismus
Lange als exotisches Randphänomen ignoriert, wird Trichoplax adhaerens heute als möglicher biologischer Modellorganismus angesehen. Insbesondere soll erforscht werden, wie sich ein Zellverband organisiert, der noch nicht als vollwertiges Epithelgewebe angesehen werden kann, wie Fortbewegung und Koordination in Abwesenheit von echtem Muskel- oder Nervengewebe funktionieren und wie sich das Fehlen einer festen Körperachse auf die Biologie des Tieres auswirkt. Auf genetischer Ebene soll untersucht werden, wie Trichoplax adhaerens Schäden an seinem Genom verhindert; insbesondere wird die Existenz spezieller DNA-Reparaturprozesse in Betracht gezogen. Die vollständige Entzifferung des Genoms soll zudem die immer noch umstrittene stammesgeschichtliche Position der Placozoa aufklären.
Neben der Grundlagenforschung könnten sich die Tiere auch zum Studium von Wundheilungs- und Regenerationsprozessen eignen; bisher unidentifizierte Stoffwechselprodukte sollen auf potentielle pharmazeutische Anwendungen hin untersucht werden. Schließlich wird Trichoplax adhaerens auch als Tiermodell zum Testen von Arzneimitteln in Betracht gezogen.
Stammesgeschichte
Die stammesgeschichtlichen Verwandtschaftsverhältnisse der Placozoa sind umstritten. Fossile Nachweise existieren nicht und sind aufgrund des weichen Körpers auch nicht zu erwarten, so dass die Stellung von Trichoplax adhaerens allein aufgrund des Vergleichs moderner Arten erschlossen werden muss.
Die klassische Zuordnung zu den Gewebelosen Tieren (Parazoa) beruht nicht auf den angenommenen Verwandtschaftsbeziehungen der Placozoa, sondern klassifiziert die Tiere stattdessen nach ihrem Organisationsgrad: Ebenso wie den Schwämmen (Porifera), mit denen sie in diesem Taxon vereinigt sind, fehlen ihnen Gewebe oder Organe; das Epitheloid wird in diesem Zusammenhang nicht als vollwertiges Gewebe angesehen. Allerdings sind dies „primitive Merkmale“, so genannte Symplesiomorphien, die auf die gemeinsame Stammart aller Tiere zurückgehen und daher nach Ansicht der heute von vielen Biologen vertretenen phylogenetischen Systematik keine evolutionäre Verwandtschaft begründen können.
Funktionsmorphologische Hypothese
 Aus einer Gallertoide entstehen durch Differenzierungsprozesse die Placozoa einerseits und die Schwämme, Nesseltiere und Rippenquallen andererseits
Aus einer Gallertoide entstehen durch Differenzierungsprozesse die Placozoa einerseits und die Schwämme, Nesseltiere und Rippenquallen andererseitsAufgrund ihres einfachen Aufbaus werden die Placozoa oft als Modellorganismen für den Übergang von den Einzellern zu den vielzelligen Tieren (Metazoa) angesehen und gelten damit als Schwestergruppe aller anderen Vielzeller:
Vielzellige Tiere (Metazoa) ├─Placozoa └─N. N. ├─Schwämme (Porifera) └─Gewebetiere (Eumetazoa)
Nach einem funktionsmorphologischen Modell stammen etwa alle Tiere von einer Gallertoide ab, einer frei im Meerwasser lebenden (pelagischen) Kugel, die von einer einlagigen begeißelten Zellschicht gebildet wird, der eine dünne, nichtzellige Trennschicht, die Basallamina, unterliegt. Das Innere der Kugel ist mit kontraktilen Faserzellen und einer gallertartigen extrazellulären Matrix gefüllt. Aus diesem vielzelligen Ausgangsstadium haben sich demnach durch zwei verschiedene Prozesse die modernen Placozoa einerseits und alle anderen Tiere andererseits gebildet:
- Einfaltungsprozesse des Epithels führten zur Ausbildung eines inneren Kanalsystems und damit zur Entwicklung einer modifizierten Gallertoide, aus der sich im folgenden die Schwämme (Porifera), Nesseltiere (Cnidaria) und Rippenquallen (Ctenophora) entwickelten.
- Andere Gallertoiden sind nach dem Modell mit der Zeit zu einer benthischen Lebensweise übergegangen, das heißt, ihr Lebensraum hat sich aus dem freien Meer auf den Gewässergrund (Benthal) verlagert. Während für frei im Wasser schwebende Tiere die Wahrscheinlichkeit, auf Beute, potentielle Sexualpartner oder Fressfeinde zu stoßen, für jede Richtung gleich groß ist, gibt es am Meeresgrund einen eindeutigen Unterschied zwischen der substratzugewandten und der substratabgewandten Seite sowie der vertikal, senkrecht zum Substrat verlaufenden Richtung. Dadurch ergibt sich auf natürliche Weise ein selektiver Vorteil für eine Abflachung des Körpers wie sie denn auch bei zahlreichen benthisch lebenden Arten zu beobachten ist. Aus einer solchen Lebensform, die nun als Placuloide bezeichnet wird, sind nun nach dem angesprochenen funktionsmorphologischen Modell sowohl die Placozoa als auch möglicherweise einige andere, nur fossil erhaltene Organismen hervorgegangen. Drei verschiedene Lebensstrategien haben demnach zu drei verschiedenen Entwicklungslinien geführt:
- Tiere, die interstitial im Sand des Meeresbodens lebten, waren demnach für die fossilen Kriechspuren verantwortlich, die als früheste Nachweise von Tieren gelten und bereits vor Anbruch der erdgeschichtlichen Epoche des Ediacariums nachweisbar sind. Normalerweise auf zweiseitig-symmetrische Würmer zurückgeführt, sieht die hier vorgestellte Hypothese auf Placuloiden zurückzuführende Tiere und damit nahe Verwandte von Trichoplax adhaerens als Urheber der Spuren an.
- Tiere, die Algen als photosynthetisch aktive Endosymbionten aufnahmen, also ihre Nährstoffe in erster Linie von ihren Symbiose-Partnern erhielten, waren demzufolge für die mysteriösen, keiner modernen Tiergruppe zugeordneten Lebewesen der Ediacara-Fauna verantwortlich, die während des Ediacariums vor Anbruch des Erdaltertums lebten.
- Tiere, die Algenrasen abweideten, waren schließlich die unmittelbaren Vorfahren der Placozoa. Die Vorteile einer amöboiden Formenvielfalt ließ demnach eine ursprünglich vorhandene Basallamina und eine gallertige extrazelluläre Matrix sekundär verlorengehen. Eine ausgeprägte Differenzierung zwischen substratzugewandter Bauch- und substratabgewandter Rückenseite führte demnach zu den noch heute nachweisbaren physiologisch unterschiedlichen Zellschichten von Trichoplax adhaerens. Diese sind demnach analog, aber nicht homolog zu Ektoderm und Entoderm, den „äußeren“ und „inneren“ Zellschichten der Gewebetiere, das heißt, die einander funktionell entsprechenden Strukturen haben nach der angegebenen Hypothese keinen gemeinsamen evolutionären Ursprung.
Falls die oben angeführte Analyse sich als richtig erweisen sollte, wäre Trichoplax adhaerens der älteste Zweig der vielzelligen Tiere und ein Relikt der Ediacara- oder sogar prä-Ediacara-Fauna. Wegen der fehlenden extrazellulären Matrix und Basallamina war das Entwicklungspotential der in ihrer ökologischen Nische sehr erfolgreichen Tiere allerdings beschränkt, was die Bradytelie genannte niedrige Evolutionsrate ihres Phänotyps, also ihrer erwachsenen Erscheinungsform, erklären würde.
Epitheliozoa-Hypothese
Funktionalmorphologische Hypothesen sind wissenschaftlich nicht unumstritten und werden häufig wegen ihres stark theoretischen und der empirischen Untersuchung nicht direkt zugänglichen Charakters abgelehnt. Die Kladistik, eine moderne Form der systematischen Forschung, stützt sich ausschließlich auf nachweisbare Merkmale moderner und fossiler Tiergruppen, um den Stammbaum einer Art oder Tiergruppe zu rekonstruieren.
Das wichtigste, auf rein morphologischen Eigenschaften beruhende Konzept sieht die Placozoa als nächste Verwandte der Gewebetiere (Eumetazoa). Das gemeinsame, als Epitheliozoa bezeichnete Taxon wird seinerseits als Schwestergruppe der Schwämme (Porifera) aufgefasst:
Vielzellige Tiere (Metazoa) ├─Schwämme └─Epitheliozoa ├─Placozoa └─Gewebetiere
Für ein solches Verhältnis sprechen in erster Linie spezielle Zell-Zell-Verbindungen, die Gürteldesmosomen, die nicht nur bei den Placozoa, sondern bei allen Tieren außerhalb der Schwämme vorkommen und dafür sorgen, dass sich Zellen zu einer lückenlosen Schicht wie dem Epitheloid der Placozoa zusammenschließen können. Auch die auf der Bauchseite auftretenden Drüsenzellen teilt Trichoplax adhaerens mit den meisten Gewebetieren. Beide Eigenschaften können als Apomorphien, also als evolutionär abgeleitete Merkmale, gelten und begründen so ein gemeinsames Taxon aller betroffenen Tiere.
Ein mögliches, von der angegebenen Hypothese inspiriertes Szenario geht davon aus, dass sich die einfach begeißelten Zellen des Epitheloids von Trichoplax adhaerens durch Reduktion des Kragens von Kragengeißelzellen (Choanozyten) der Schwämme herausgebildet haben, als sich die Vorfahren der Placozoa von einer filtrierenden Lebensweise abwandten. Das Epitheloid gilt dann als Vorläufer des echten Epithelgewebes der Gewebetiere.
Anders als in dem oben angesprochenen funktionalmorphologisch begründeten Modell gelten im Epitheliozoa-Konzept die bauch- und rückseitigen Zellschichten der Placozoa als Homologien zu Endoderm und Ektoderm, den beiden grundlegenden embryonalen Zellschichten der Gewebetiere – aus Endoderm entwickelt sich etwa die verdauungsaktive Gastrodermis der Nesseltiere (Cnidaria) oder das Darmepithel der zweiseitig symmetrischen Bilateria, während Ektoderm unter anderem der Vorläufer der äußeren Hautschichten (Epidermis) ist. Der bei den Placozoa von einem Fasersynzytium durchsetzte Innenraum entspräche dann dem Bindegewebe der anderen Tiere. Ob die im Synzytium gespeicherten Kalzium-Ionen in einem Zusammenhang mit den Kalkskeletten vieler Nesseltiere stehen, ist ungewiss.
Eumetazoa-Hypothese
Eine dritte, in erster Linie molekulargenetisch begründete Hypothese sieht die Placozoa als stark vereinfachte Gewebetiere. Demnach stammt Trichoplax adhaerens von wesentlich komplexer aufgebauten Tieren ab, die bereits über Muskeln und Nervengewebe verfügten. Beide Gewebetypen wie auch die Basallamina des Epithels sind demzufolge erst durch radikale sekundäre Vereinfachung verloren gegangen.
Verschiedene Studien kommen dabei derzeit zu unterschiedlichen Ergebnissen hinsichtlich der genauen Schwestergruppe: Mal gelten die Placozoa als nächste Verwandte der Nesseltiere (Cnidaria), mal als Schwestergruppe der Rippenquallen und gelegentlich werden sie sogar direkt neben die zweiseitig-symmetrischen Bilateria gestellt:
Vielzellige Tiere (Metazoa) ├─Schwämme (Porifera) └─Gewebetiere (Eumetazoa) ├─Rippenquallen (Ctenophora) └─N. N. ├─Nesseltiere (Cnidaria) └─N. N. ├─Placozoa └─BilateriaLediglich eine ebenfalls vorgeschlagene Einordnung in die Nesseltiere lässt sich derzeit mit hoher Wahrscheinlichkeit ausschließen.
Kritisch wird gegen das vorgeschlagene Szenario eingewandt, dass es morphologische Merkmale der Tiere vollkommen unberücksichtigt lässt. Eine so extreme Vereinfachung, wie sie dem Modell gemäß für die Placozoa postuliert werden müsste, ist zudem nur von parasitisch lebenden Organismen bekannt, ließe sich aber für eine frei lebende Tierart wie Trichoplax adhaerens nur schwer funktional erklären.
Systematik
Derzeit wird nur eine einzige Art, Trichoplax adhaerens, als Bestandteil der Placozoa anerkannt. Allerdings wurde 1893 durch den Italiener Francesco Saverio Monticelli eine weitere Art mit dem Namen Treptoplax reptans beschrieben, die er in den Gewässern um Neapel fand. Sie gilt jedoch seit 1896 als verschollen; ihre Existenz wird heute von den meisten Zoologen angezweifelt.
Weil zwischen Vertretern von Trichoplax adhaerens oft große genetische Unterschiede bestehen, die bei anderen Organismengruppen zu einer Einteilung in unterschiedliche Gattungen führen würden, ist derzeit unklar, ob die nach morphologischen Kriterien einzige Art nicht tatsächlich einer Gruppe kryptischer, also äußerlich nicht unterscheidbarer Arten entspricht. Das Vorkommen der genetischen Varianten ist nicht geographisch bedingt: Einige Varianten sind in verschiedenen Regionen nachgewiesen (z. B. Pazifik, Karibik und Rotes Meer). Gleichzeitig lassen sich sehr unterschiedliche genetische Varianten aus demselben Lebensraum isolieren.
Einzelnachweise
- ↑ Rüdiger Wehner, Walter Gehring: Zoologie. 24. Auflage, Thieme, Stuttgart, Juni 2007 S. 696
Literatur
- Edward E. Ruppert, R. S. Fox, R. D. Barnes: Invertebrate Zoology – a functional evolutionary approach. Kapitel 5. Brooks/Cole, London 2004 (7.Aufl.), S. 94, ISBN 0-03-025982-7
- Richard C. Brusca, G. J. Brusca: Invertebrates. Kapitel 7. Sinauer Associates, Sunderland Mass 2002 (2. Aufl.), S. 210, ISBN 0878930973
Wissenschaftliche Literatur
- V. J. Birstein: On the Karyotype of „Trichoplax sp.“ (Placozoa). in: Biologisches Zentralblatt. Fischer, Jena – Stuttgart 108.1989, S. 63, ISSN 0006-3304
- K. G. Grell, A. Ruthmann: Placozoa. in: F. W. Harrison, J. A. Westfall (Hrsg.): Microscopic Anatomy of Invertebrates. Bd 2. Wiley-Liss, New York 1991, S. 13, ISBN 0471562246
- W. Jakob, S. Sagasser, S. Dellaporta, P. Holland, K. Kuhn, B. Schierwater: The Trox-2 Hox/Para Hox gene of „Trichoplax“ (Placozoa) marks an epithelial boundary. in: Development Genes and Evolution. Springer, Berlin 214.2004, S. 170, ISSN 0949-944X
- Y. K. Maruyama: Occurrence in the Field of a Long-Term, Year-Round Stable Population of Placozoa. in: Biological Bulletin. Laboratory, Woods Hole Mass 206.2004,1, S. 55, ISSN 0006-3185
- T. Syed, B. Schierwater: The evolution of the Placozoa – A new morphological model. in: Senckenbergiana lethaea. Schweizerbart, Stuttgart 82.2002,1, S. 315, ISSN 0037-2110
- P. Schubert: „Trichoplax adhaerens“ (Phylum Placozoa) has cells that react with antibodies against the neuropeptide RFAmide. in: Acta Zoologica. Blackwell Science, Oxford 74.1993,2, S. 115, ISSN 0001-7272
- T. Ueda, S. Koga, Y. K. Marayama: Dynamic patterns in the locomotion and feeding behaviour by the placozoan „Trichoplax adhaerens“. in: BioSystems. North-Holland Publ., Amsterdam 54.1999, S. 65, ISSN 0303-2647
- O. Voigt, A.G. Collins, V. Buchsbaum Pearse, J.S. Pearse, A. Ender, H. Hadrys, B. Schierwater: Placozoa – no longer a phylum of one. in: Current Biology. Cell Press, Cambridge Mass 14.2004, 22, S. R944, ISSN 0960-9822
Erstbeschreibungen
- Placozoa
- K. G. Grell: Trichoplax adhaerens, F. E. Schulze und die Entstehung der Metazoen. in: Naturwissenschaftliche Rundschau. Wiss. Verl.-Ges., Stuttgart 24.1971, S. 160, ISSN 0028-1050
- Treptoplax reptans
- F. S. Monticelli: Treptoplax reptans n. g., n. s. in: Rendiconti / Accademia Nazionale dei Lincei, Roma, Classe di Scienze Fisiche, Matematiche e Naturali. Rom 2.1893,5, S. 39, ISSN 0001-4435
- Trichoplax adhaerens
- F. E. Schulze: Trichoplax adhaerens n. g., n. s. in: Zoologischer Anzeiger. Elsevier, Amsterdam-Jena 6.1883, S. 92, ISSN 0044-5231
Weblinks



Wikimedia Foundation.