- Blattgrünkörper
-

Die Chloroplasten (von griech. χλωρός chlōrós „grün“) sind Organellen der Zellen von Grünalgen und höheren Pflanzen, die Photosynthese betreiben. Bei höheren Pflanzen können aus den photosynthetisch aktiven Chloroplasten durch Differenzierung Chromoplasten, Leukoplasten (Amyloplasten, Etioplasten) und Gerontoplasten hervorgehen (zusammengefasst als Plastiden).
Inhaltsverzeichnis
Bau der Chloroplasten
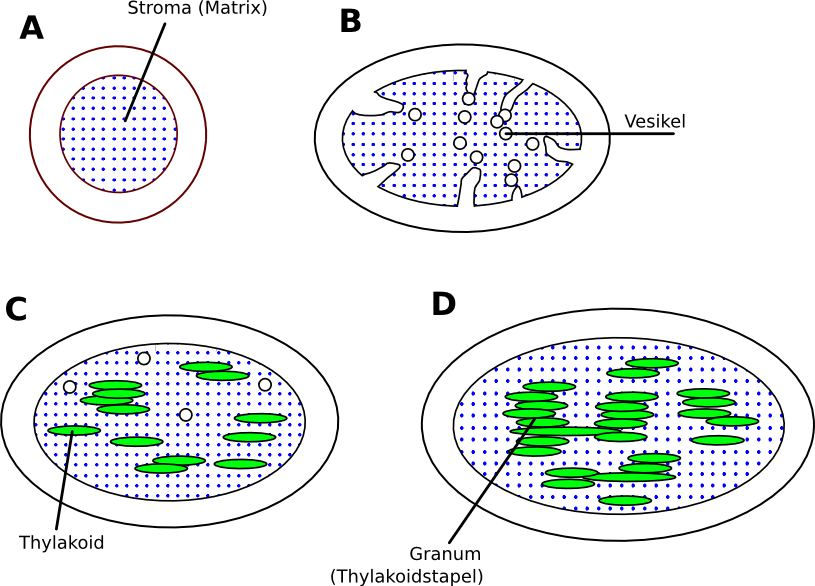
 Lichtinduzierte Entwicklung eines Chloroplasten aus einem Proplastiden:
Lichtinduzierte Entwicklung eines Chloroplasten aus einem Proplastiden:
A: Der Proplastid aus unbelichteten Pflanzen besteht lediglich aus einer Innen- und einer Außenmembran
B: Licht induziert die Synthese von Chlorophyll, Phospholipiden und Thylakoid-Proteinen. Von der Innenmembran schnüren sich Vesikel in den Matrixraum ab
C: Während der Vergrößerung der Proplastiden bilden sich einzelne Thylakoide durch Fusion von Vesikeln. Durch ein Protein der Lichtsammelkomplexe kommt es dann zur Verklebung (Stapelung) von Thylakoiden.
D: Durch Ausbau der Membransysteme, wobei die Grana-Thylakoidstapel durch (ungestapelte) Stroma-Thylakoide miteinander verbunden werden, entsteht der Chloroplast Modell eines Chloroplasten (im Überseemuseum Bremen)
Modell eines Chloroplasten (im Überseemuseum Bremen)Ähnlich wie Mitochondrien besitzen Chloroplasten sowohl eine eigene DNA als auch eigene Ribosomen und zwei Biomembranen als Hülle. In ihrem Innern befindet sich als plasmatische Phase das Stroma. Das Stroma wiederum wird von Thylakoidmembranen durchzogen - Abkömmlingen der Innenmembran. Mit Ausnahme vieler phototropher Protisten sind in den Chloroplasten der höheren Phototrophen an etlichen Stellen flache, runde Ausstülpungen dieser Membranen „geldrollenartig“ übereinandergelagert. Einen solchen Thylakoidstapel nennt man Granum (pl. Grana). In die Membranen der Thylakoide sind verschiedene Pigmente eingelagert, vor allem der grüne Farbstoff Chlorophyll. Besonders viel davon findet sich in den Membranen der Grana, weshalb diese intensiv grün gefärbt erscheinen. Die Pigmente können Licht bestimmter Wellenlängen absorbieren und die aufgenommene Energie wird zur Produktion von ATP aus ADP und Phosphat genutzt (siehe Phototrophie). ATP dient als Energieüberträger zum Aufbau von Glucose bzw. Stärke aus CO2 und Wasser (siehe Photosynthese, Calvin-Zyklus, Gluconeogenese).
Aufbau eines Protonengradienten
Die Biogenese dieser drei Membransysteme erklärt die Tatsache, dass der Protonengradient bei Chloroplasten über die Thylakoidmembran aufgebaut wird (das Thylakoid-Innere weist ein saures Milieu auf), während bei Mitochondrien der Intermembranraum (Bereich zwischen Innen-und Außenmembran) mit Protonen beladen wird. Analog ist die ATP-Synthase (alias FoF1-ATPase) bei Chloroplasten ein in die Thylakoidmembran eingebettetes Enzym (CF1-Teil ragt ins Stroma), bei Mitochondrien ein Bestandteil der Innenmembran (F1-Teil der Matrix zugewandt). Bei beiden Systemen wird ATP also an die Matrix/das Stroma abgegeben. Im Gegentausch zu ADP kann es in das Cytosol der Zelle gelangen.
Herkunft der Chloroplasten - Endosymbiontentheorie
Der strukturelle Aufbau eines Chloroplasten gleicht dem eines Cyanobakteriums (Blaualge). Er besitzt wie dieses ein ringförmig strukturiertes DNA-Genom und eine eigene Proteinsynthese. Der Aufbau der hierbei aktiven Ribosomen ist identisch mit denen in Cyanobakterien. Ein weiteres Argument ist die eigenständige „Vermehrung“ der Chloroplasten ohne eine strukturelle Koppelung an die Zellteilung der umgebenden Eukaryontenzelle.
Dies hat zu der Endosymbiontentheorie geführt, die besagt, dass Chloroplasten sich als endosymbiotische Cyanobakterien und Mitochondrien als endosymbiotische Bakterien entwickelt haben (s. auch Andreas Franz Wilhelm Schimper, Konstantin Sergejewitsch Mereschkowski, Lynn Margulis).
Im Zuge der Evolution und Eingliederung des cyanobakteriellen Vorläufers der Chloroplasten in die „Wirtszelle“ kam es zu diversen Anpassungen. Hierzu zählt unter anderem die Anpassung des Chloroplastengenoms. Die Größe des Genoms sank von ca. 3,5 Mio. Basen auf 120 - 160 kb. Zusätzlich bildete sich eine vierteilige Struktur des Genoms heraus, die in den meisten photosynthetischen Organismen zu finden ist. Hierbei handelt es sich um zwei gegenläufige Kopien, die von einmaligen Stücken unterbrochen werden, wobei die Größen der drei unterschiedlichen DNA.Regionen speziesabhängig bis hin zum vollständigen Verlust der kopierten Regionen variieren kann. Die Verkleinerung des Genoms ging mit Verlust genetischer Information und Transfer in den Zellkern einher. Gleichzeitig entwickelte sich eine komplexe Maschinerie zum Import von Proteinen aus dem Zytosol in den Chloroplasten (TIC, TOC). So findet man ca. 2000 Proteine im Chloroplasten, wobei nur 100 Gene im Genom zu finden sind. Die Gene kodieren für Produkte, die sich grob in zwei Kategorien einteilen lassen: Aufrechterhaltung des genetischen Apparats (DNA-Polymerase, tRNAs und rRNAs) und Aufrechterhaltung der photosynthetischen Kapazität (Photosystemkomponenten und weitere Proteine). Bisher ist nicht vollständig geklärt, wie die Synchronisation der Expression zwischen Kern und Chloroplasten erfolgt. Dies ist notwendig, da in allen Proteinkomplexen im Chloroplasten plastidär und nukleär kodierte Produkte zusammengesetzt werden.
Lange Zeit war unbekannt, wie Chloroplasten sich teilen und ihre Form verändern. Heute weiß man, dass auch Bakterien ein Zytoskelett besitzen, dessen Proteine evolutionäre Vorläufer des eukaryotischen Zytoskeletts sind. Aus Versuchen am Laubmoos Physcomitrella patens ist bekannt, dass die FtsZ-Proteine, die Tubulin-Homologen, nicht nur die Teilung der Chloroplasten bewirken [1], sondern auch ein komplexes Netzwerk in den Chloroplasten ausbilden können. Da diese Netzwerke stark an das Zytoskelett erinnern, prägte Ralf Reski 2000 den Begriff „Plastoskelett“ für diese Struktur und postulierte, dass sie ähnlich komplexe Funktionen in den Plastiden ausfüllt, wie das Zytoskelett für die gesamte Zelle. [2][3]
Siehe auch
- Chemiosmotische Kopplung
- Endosymbiontentheorie (zum Ursprung der Chloroplasten)
Einzelnachweise
- ↑ Strepp, R. et al. (1998): Plant nuclear gene knockout reveals a role in plastid division for the homolog of the bacterial cell division protein FtsZ, an ancestral tubulin. In: Proc. Natl. Acad. Sci. USA 95(8); 4368–4373; PMID 9539743; PDF (freier Volltextzugriff, engl.)
- ↑ Reski, R. (2002): Rings and networks: the amazing complexity of FtsZ in chloroplasts. In: Trends in Plant Science 7(3); 103–105; PMID 11906832; doi:10.1016/S1360-1385(02)02232-X
- ↑ Kiessling, J. et al. (2000): Visualization of a cytoskeleton-like FtsZ network in chloroplasts. In: Journal of Cell Biology 151(4); 945–950; PMID 11076976; PDF (freier Volltextzugriff, engl.)
Literatur
- D. von Wettstein (1959): The effect of genetic factors on the submicroscopic structures of the chloroplast, J. Ultrastruct. Res. 3, 235-239.
Weblinks
- Aufbau von Chloroplasten - Schaubild
- http://www.zytologie-online.net/photosynthese.php - [ Chloroplast und Photosynthese / Energiegewinnung bei Pflanzen] - Grafik / Animation


Wikimedia Foundation.