- Cardiovaskuläres System
-
Der Blutkreislauf ist das Strömungssystem des Blutes, das vom Herzen und, wenn vorhanden, einem Netz aus Blutgefäßen (kardiovaskuläres System) gebildet wird.
Er sichert das Überleben des Organismus, indem er den Stoffwechsel jeder einzelnen Körperzelle versorgt und die chemischen und physiologischen Eigenschaften der Körperflüssigkeiten aufrecht erhält. Zum einen transportiert das Blut Sauerstoff aus den Lungen zu den Zellen und Kohlendioxid in entgegengesetzter Richtung (siehe auch Atmung). Zum anderen werden aus der Verdauung gewonnene Nährstoffe wie Fette, Zucker oder Eiweiße aus dem Verdauungstrakt in die einzelnen Gewebe transportiert, um dort je nach Bedarf verbraucht, weiterverarbeitet oder gespeichert zu werden. Die entstandenen Stoffwechsel- oder Abfallprodukte (zum Beispiel Harnstoff oder Harnsäure) werden dann in anderes Gewebe oder zu den Ausscheidungsorganen (Nieren und Dickdarm) transportiert. Außerdem verteilt das Blut auch Botenstoffe wie zum Beispiel Hormone, Zellen der Körperabwehr und Teile des Gerinnungssystems innerhalb des Körpers.
Inhaltsverzeichnis
Formen
Bedingt durch die Entwicklungsgeschichte der Tiere gibt es unterschiedlich ausgeprägte Arten von Kreisläufen:
- Manche Tiere, wie z. B. die Plattwürmer, besitzen keinerlei Kreislaufsystem. Ihr Mund führt direkt in ein verästeltes Verdauungssystem, aus dem Nährstoffe wegen der Flachheit des Wurmes direkt in alle Zellen diffundieren können. Sauerstoff diffundiert aus dem Wasser in die Zellen.
- Bei einer Reihe von wirbellosen Tieren wie den Gliederfüßern und Weichtieren findet man einen offenen Kreislauf, bei dem das Blut in den Körperhöhlen kreist. Hier wird die Körperflüssigkeit, die man als Hämolymphe bezeichnet, vom Herzen in kurze Gefäße und von dort in alle Körperhöhlen gepumpt, bis es schließlich ins Herz zurückfließt. Die Hämolymphe fließt dabei langsam und mit geringem Druck.
- Bei Ringelwürmern wie dem Regenwurm existiert ein geschlossener Kreislauf, bei dem die blutähnliche Körperflüssigkeit durch kontraktile Gefäße in Bewegung gehalten wird.
- Auch Wirbeltiere haben einen geschlossenen Kreislauf. Hier fließt das Blut, welches sich von der Hämolymphe unterscheidet, immer durch ein geschlossenes Netz aus Blutgefäßen, das alle lebenden Zellen erreicht. Herz, Blutgefäße und Blut bilden dabei das so genannte Herz-Kreislauf-System. Dieses teilt sich entwicklungsgeschichtlich noch einmal auf und unterscheidet die wechselwarmen von den gleichwarmen Wirbeltieren.
- Bei den wechselwarmen (poikilothermen) Tieren, also Fischen, Amphibien und Reptilien, findet eine Vermischung von sauerstoffreichem und sauerstoffarmem Blut im Herzen statt, weil nur eine oder zwei unvollständig getrennte Herzkammern vorliegen.
- Bei den gleichwarmen (homoiothermen)Tieren, den Vögeln und Säugetieren, besteht das Herz aus zwei Vorhöfen und zwei Kammern, so dass eine vollständige Trennung von sauerstoffreichem und sauerstoffarmen Blut besteht.
Allgemeines zum geschlossenen Blutkreislauf
Aufbau
Der Blutkreislauf besteht aus dem Herzen und den Blutgefäßen. Blutgefäße, die zum Herzen führen, werden als Venen bezeichnet, diejenigen, die vom Herz wegführen, als Arterien. Je weiter die Blutgefäße vom Herzen entfernt sind, umso verzweigter werden sie, und umso kleiner wird auch ihr Durchmesser. Arterien werden zuerst zu Arteriolen und diese zu Kapillaren, welche das Gewebe versorgen. Diese führen wiederum zusammen und bilden die postkapillaren Venolen, die zu Venen werden.
Blutgefäße
Blutgefäße werden auf Grund ihres Aufbaus und ihrer Funktion in mehrere Arten unterteilt. Die Arterien transportieren das Blut unter hohem Druck und mit hoher Fließgeschwindigkeit, deswegen besitzen sie eine dicke Gefäßwand. Durch sie gelangt das Blut aus dem Herzen in die verschiedenen Gewebe. Von den Arterien gehen die Arteriolen ab, sie dienen als Kontrollventile, und haben deswegen starke muskuläre Wände, die die Gefäße verschließen (Vasokonstriktion) oder weiten (Vasodilation) können. Sie verzweigen sich weiter zu den Kapillaren, die den Austausch von Flüssigkeiten, Nährstoffen, Elektrolyten, Hormonen und anderen Stoffen zwischen Blut und Gewebe vornehmen und deswegen mit einer dünnen Gefäßwand (nur Endothel) ausgestattet sind, die für geringmolekulare Stoffe permeabel (durchlässig) ist. In einigen Organen (Leber, Milz) sind die Kapillaren erweitert, dann spricht man von Sinusoiden.
Venolen haben nur eine dünne Gefäßwand. Sie sammeln das Blut aus den Kapillaren, um es wieder den Venen zuzuführen, die es von der Peripherie zurück zum Herzen transportieren. Weiterhin dienen sie als Blutspeicher. Sie haben dünne, muskuläre Wände, die das Weiten oder Verschließen der Gefäße erlauben. Ein Teil der Flüssigkeit tritt im Kapillargebiet aus den Gefäßen aus und wird über Lymphgefäße abtransportiert. Die großen Lymphsammelstämme münden nahe dem Herzen wieder in das Venensystem.
Benachbarte Blutgefäße mit gleichem Zielgebiet werden als Kollaterale bezeichnet. In fast allen Körperregionen gibt es Verbindungen zwischen diesen benachbarten Blutgefäßen, so genannte Anastomosen. Diese sorgen dafür, dass bei einer Verlegung (etwa einer Thrombose) oder Verletzung eines Blutgefäßes, die Versorgung durch das Nachbargefäß übernommen werden kann. Arterien, die keine Anastomosen aufweisen, nennt man Endarterien. Kommt es zu einer Verlegung einer Endarterie, so wird der entsprechende Gewebsabschnitt nicht mehr mit Blut versorgt und stirbt ab (Infarkt). Die Anastomosen können aber auch zu schwach sein, um eine vollständige Kompensation eines Ausfalls zu ermöglichen. In diesem Fall spricht man von funktionellen Endarterien. Eine Verstopfung oder Verletzung dieser Arterien führt zu einer Minderdurchblutung (Ischämie).
Aufgaben und Funktionen
Blut erfüllt im Körper verschiedene Aufgaben. Es transportiert sauerstoffreiches Blut aus den Lungen zum Gewebe und mit Kohlenstoffdioxid angereichertes Blut zurück. Weiterhin wird das Gewebe mit Nährstoffen aus dem Verdauungstrakt versorgt und von entstandenen Stoffwechsel- und Abfallprodukten befreit, die zu den Ausscheidungsorganen (Niere und Darm) transportiert werden. Blut dient zudem als wichtiges Medium für den Transport von Hormonen zwischen einzelnen Organsystemen und Komponenten der Immunabwehr und der Blutgerinnung zu Orten im Körper an denen sie gebraucht werden.
Der Blutkreislauf dient demzufolge letztendlich dazu, dem Blut zu ermöglichen, sich durch den gesamten Körper zu bewegen. Weiterhin spielt der Blutkreislauf eine wichtige Rolle bei der Thermoregulation. Über den Grad der Durchblutung der Haut wird die Wärmeabgabe über die Körperoberfläche reguliert.Blutkreislauf der wechselwarmen Wirbeltiere
Fische
 Schematische Darstellung des Blutkreislaufs der Fische
Schematische Darstellung des Blutkreislaufs der Fische
Rot = Sauerstoffreiches Blut,
Blau = Sauerstoffarmes BlutDas Herz der Fische ist das am einfachsten gebaute unter den Wirbeltieren, denn es besteht nur aus einem dünnwandigem Vorhof und einer dickwandigen, muskulösen Kammer. Zwischen den beiden befindet sich eine einzelne Klappe, die einen Rückstrom des Blutes verhindert. Ebenso wie das Herz ist auch der Blutkreislauf selbst relativ einfach strukturiert. Das venöse Blut wird aus dem Herzen in die Kiemen gepumpt (Kiemenkreislauf), in denen es mit Sauerstoff aus dem Wasser angereichert wird. Anschließend wird das sauerstoffreiche Blut in den Körperkreislauf weitertransportiert. In den Kapillaren gibt es den Sauerstoff ab und nimmt dafür Kohlendioxid auf. Neben dem Herz nimmt auch die Muskulatur der Kiemen am Pumpvorgang teil. Der Nachteil dieser Konstruktion ist, dass der Blutdruck im Kapillarnetz des Kiemenkreislaufs stark abfällt, der Blutstrom durch den Körper also relativ langsam erfolgt. Zudem haben Fische ein geringes Blutvolumen. Es macht weniger als ein Zehntel des Körpergewichts aus. Außerdem liegt der Sauerstoffgehalt im Blut eines Fisches weit unter dem des menschlichen Blutes.
Amphibien
Doppelter Kreislauf
 Schematische Darstellung des Blutkreislaufs der Amphibien
Schematische Darstellung des Blutkreislaufs der Amphibien
Rot = Sauerstoffreiches Blut,
Blau = Sauerstoffarmes Blut
Rosa = MischblutBei den Amphibien (Lurche) besteht das Herz aus einer Kammer und zwei Vorhöfen. Der Gasaustausch findet sowohl in der Lunge als auch in der Haut statt. Die beiden Kreisläufe der Amphibien werden daher als Lungen-Haut-Kreislauf und Körperkreislauf bezeichnet. Da sie, im Gegensatz zu Fischen, nicht hintereinander geschaltet sind, spricht man von einem doppelten Kreislauf.
Der linke Vorhof empfängt mit Sauerstoff angereichertes Blut aus der Lunge und der Haut, der rechte Vorhof sauerstoffarmes Blut aus dem Körper. Beide Vorhöfe pumpen das Blut in die einheitliche Kammer. Diese Kammer besitzt einen Ausflusstrakt (Truncus oder Conus arteriosus), der sich in jeweils einen Stamm für die beiden Kreisläufe teilt. Eine leistenartige Erhebung im Ventrikel und im Lumen des Ausflusstrakts sorgt dafür, dass das Blut relativ „sortenrein“ durch das Herz fließt, sauerstoffarmes und -reiches Blut sich also nur wenig vermischen. Das sauerstoffreichere Blut wird zum überwiegenden Teil in die Halsschlagadern und die Aorta gepumpt, während das sauerstoffärmere Blut in die Lungen-Haut-Arterie gelenkt wird. Wie Reptilien und Vögel besitzen die Amphibien bereits einen Nierenpfortaderkreislauf.
Entwicklung
Amphibien haben ursprünglich vier paarige Kiemenbogenarterien, die zu beiden Seiten aus der Aorta entspringen. Bei ausgewachsenen Lurchen entwickelt sich die erste zur Arteria carotis, die den Kopf versorgt. Die Arterien des zweiten Bogens vereinigen sich zur Aorta descendens, der absteigenden Aorta. Die dritte Kiemenbogenarterie bildet sich zurück, und aus den vierten entwickelt sich der paarige Aortenbogen.
Reptilien
 Schematische Darstellung des Blutkreislaufs der Reptilien
Schematische Darstellung des Blutkreislaufs der Reptilien
Rot = Sauerstoffreiches Blut,
Blau = Sauerstoffarmes Blut
Rosa = MischblutDie zu den Reptilien zusammengefassten Taxa besitzen ein Herz, das ebenso aus zwei Vorhöfen und einer Kammer besteht. Diese ist jedoch nahezu vollständig durch eine Scheidewand in zwei Hälften geteilt. Aus dem Körper strömt sauerstoffarmes Blut in den rechten Vorhof, aus den Lungen mit Sauerstoff angereichertes Blut fließt in den linken Vorhof. Beide Vorhöfe pumpen das Blut in die Herzkammer, aus der drei Schlagadern abgehen. In der rechten fließt sauerstoffarmes Blut zur Lunge, in der linken sauerstoffreiches Blut zum Kopf und in den Körper. Da die Trennung der Herzkammer jedoch nicht vollständig ist, kommt es zur Bildung von Mischblut (circa 10 bis 40 Prozent). Dieses fließt durch die mittlere Schlagader in den Körper.
Eine Besonderheit stellen die Krokodile dar, bei ihnen sind die beiden Herzkammern komplett getrennt. Zwischen der linken und der rechten Schlagader besteht bei ihnen mit dem Foramen Panizzae eine Verbindung. Dabei entspringt die linke Aorta an der rechten Herzkammer und die rechte an der linken. Durch das Fenster vermischt sich das sauerstoffreiche Blut der rechten Kammer mit dem sauerstoffarmen der linken Kammer im Bereich der rechten Aorta, so dass Mischblut in den Körperkreislauf geführt wird und dabei vor allem in die peripheren Bereiche des Körpers gelangt. Zugleich fördert die linke Aorta sauerstoffreiches Blut in den Körper und vor allem in den Kopf des Tieres. Beim Tauchvorgang schließt sich das Foramen Panizzae vollständig, so dass die rechte Aorta nur noch mit sauerstoffarmem Blut versorgt wird, der Kopf jedoch weiterhin sauerstoffreiches Blut bekommt.
Auch bei den Dinosauriern lag vermutlich eine vollständige Trennung der Herzkammern vor, was sie zu quasi-gleichwarmen Tieren machen würde und somit deren langes Überleben erklären könnte (s. Thermoregulation). Dies ergibt sich aufgrund ihrer Position im Stammbaum zwischen den Krokodilen und den Vögeln, die beide eine durchgängige Trennwand im Herzen besitzen.
Blutkreislauf der gleichwarmen Wirbeltiere
Anatomie
 Schematischer Aufbau eines doppelten Blutkreislaufs.
Schematischer Aufbau eines doppelten Blutkreislaufs.
Rot = sauerstoffreiches Blut
Blau = sauerstoffarmes BlutIm Gegensatz zu den wechselwarmen Tieren ist das Herz der gleichwarmen Tiere, und damit auch das des Menschen, vollständig in zwei Kammern geteilt. Deshalb kann es als in zwei Hälften geteilt betrachtet werden, obwohl es sich im gesamten um ein einziges Organ handelt. Jede dieser Hälften besteht aus einem Vorhof und einer Kammer, die jeweils als Einheit arbeiten. Während die rechte Herzhälfte das Blut durch den Lungenkreislauf pumpt, der das Blut mit Sauerstoff anreichert, pumpt die linke Herzhälfte das Blut durch den Körperkreislauf, um die Organe mit Nährstoffen und Sauerstoff zu versorgen.
Diese beiden Kreisläufe sind in Reihe geschaltet, so dass das gesamte Blut immer durch den Lungenkreislauf fließen muss. Im Unterschied dazu sind die Organe im Körperkreislauf parallel geschaltet.
Die Existenz zweier Blutkreisläufe (Körper- und Lungenkreislauf) hat wichtige Vorteile:
- Der Druck kann in beiden Kreisläufen unterschiedlich sein. Im Lungenkreislauf ist er erheblich niedriger, so dass eine geringere Wanddicke in den Lungen für einen besseren Gasaustausch möglich ist.
- Die Lunge mit ihren Kapillaren funktioniert als Filter gegen Blutgerinnsel (Thromben) u. ä., bevor das Blut von der linken Herzseite u. a. zum Gehirn gepumpt wird. Die Lunge hat dazu thrombenlösende Eigenschaften.
Im Lungenkreislauf verlässt das Blut die rechte Herzkammer über den Lungenstamm (lat. Truncus pulmonalis) in Richtung der Lungen, wo es mit Sauerstoff angereichert wird. Dann wird es von der Lungenvene (lat. Vena pulmonalis ) in den linken Herzvorhof gepumpt. Vom linken Vorhof gelangt es in die linke Kammer, von wo aus es durch die Aorta in den Körperkreislauf gepumpt wird. Während bei den Säugern die Aorta auf der linken Körperseite verläuft, liegt sie bei Vögeln auf der rechten. Nach der Versorgung der Organe kehrt das nun mit Kohlendioxid angereicherte Blut durch die obere bzw. die untere Hohlvene in den rechten Vorhof zurück. Mit dem Übergang vom rechten Vorhof in die rechte Kammer beginnt der Kreislauf von neuem.
Eine Besonderheit stellt das Pfortadersystem dar. Blut, das von den Organen des Verdauungstrakts kommt, wird in der Pfortader gesammelt und gelangt in die Leber, wo die aufgenommenen Nährstoffe verwertet werden. Auch die Hirnanhangsdrüse (Hypophyse) hat ein Pfortadersystem. Vögel und Reptilien haben zudem eine Nierenpfortader.
Blutdruck und -volumen
Man unterscheidet zwischen dem so genannten Niederdruck- und dem Hochdrucksystem. Zum Niederdrucksystem gehören die Arteriolen, Kapillaren und Venen des Körperkreislaufs, das rechte Herz und die Gefäße des Lungenkreislaufs. Das Hochdrucksystem enthält die Arterien des Körperkreislaufs.
Die Hauptaufgabe des Niederdrucksystems ist seine Blutspeicherfunktion, denn 80 Prozent des im Körper zirkulierenden Blutes (etwa sieben Prozent der fettfreien Körpermasse, beim Menschen circa vier bis fünf Liter) findet man dort. Diese Funktion wird durch die hohe Dehnbarkeit und die große Kapazität der Gefäße begünstigt. Im Falle eines Blutverlustes kann das Volumen durch Verengung (Vasokonstriktion) der Venen bis zu einem gewissen Grad ausgeglichen werden. Im umgekehrten Fall, der zum Beispiel bei Bluttransfusionen auftritt, ändert sich hauptsächlich das Volumen des Niederdrucksystems. Deshalb ist im Normalfall der zentrale Venendruck (Normalwert etwa drei bis neun mmHg) ein guter Indikator für das Blutvolumen. Im Gegensatz dazu ist die Hauptaufgabe des Hochdrucksystems die Versorgung der Organe.
Der Blutdruck ist im Verlauf des Systems großen Änderungen unterworfen. Beträgt er noch in der Aorta und den großen Arterien ca. 100 mmHg, fällt er in den Arterienästen auf 40 mmHg ab, und beträgt in den Kapillaren nur noch 25 mmHg. In den Venolen beträgt er 20 mmHg, in den Pfortadern letztendlich nur noch drei mmHg. Im Lungenkreislauf schwankt der Druck zwischen 15 und 20 mmHg in der Arteria pulmonalis und zwischen 2 und 5 mmHg in der Vena pulmonalis.
Spricht man umgangssprachlich vom Blutdruck, so meint man den Blutdruck der Arterien im Körperkreislauf. Dieser schwankt zwischen Systole (der Auswurfphase des Herzens) und Diastole (der Füllungsphase), und wird als Doppelwert dieser beiden Phasen angegeben, wobei zuerst der systolische und dann der diastolische Wert genannt wird. Durchschnittlich liegen diese Werte für die Diastole zwischen 60 und 90 mmHg und für die Systole zwischen 100 und 140 mmHg. Der Unterschied zwischen dem systolischen und dem diastolischen Blutdruck wird als Blutdruckamplitude bezeichnet.
Blutfluss
Trotz der großen Druckunterschiede zwischen Systole und Diastole fließt das Blut relativ gleichmäßig durch den Körper. Dies liegt an der so genannten Windkesselfunktion der Aorta und der großen Arterien. Während der Systole dehnt sich die Gefäßwand aus und nimmt so einen Teil des ausgeworfenen Blutes auf, und gibt ihn in der Diastole, in der kein Blut aus dem Herzen austritt, wieder ab. Diese Volumendehnbarkeit (Compliance) wandelt also das stoßweise austretende Blut in einen gleichmäßigen Strom um. Würde der Druck nicht durch die elastischen Gefäße gespeichert werden können, so würde der Druck in der Aorta wesentlich dramatischer schwanken. Interessanterweise würde im zeitlichen Mittel aber wesentlich weniger Blut durch die Gefäße strömen, da viel Strömungsenergie für das ständige Beschleunigen des Blutes aufgezehrt würde.
Die Druckwelle bewegt sich beim jungen, erwachsenen Menschen mit etwa 6 Meter pro Sekunde, beim alten Menschen verdoppelt sich die Geschwindigkeit. Da mit zunehmendem Lebensalter die Gefäßwände immer unelastischer werden, vermindert sich der Druckspeichereffekt mit dem Lebensalter immer mehr und der Volumenstrom reduziert sich.
Während der Blutfluss in den Arterien allein von der Pumpkraft des Herzens realisiert wird, spielen bei Venen verschiedene Faktoren eine Rolle. Zu einem gewissen Grad wirkt die Pumpkraft über das Kapillarbett hinaus auch auf die Venen (sog. vis a tergo, „Kraft von hinten“). In den Venen wird das Blut vor allem schubweise über von außen wirkende Kräfte zurück zum Herz transportiert. Zu diesem Zweck befinden sich in ihrem Inneren Venenklappen. Die äußeren Kräfte sind vor allem die Kontraktionen umliegender Skelettmuskeln, bei den großen Venen im Körperinneren die Druckschwankungen durch die Atmung (Erweiterung der Venen durch den Unterdruck bei der Inspiration). Die Venenklappen verhindern, dass in den Pausen dieser äußeren Massagewirkung das Blut nicht wieder der Schwerkraft folgend zurück fließt. Der Ansaugdruck durch die Erweiterung der Vorhöfe des Herzens spielt nur bei den herznahen großen Venen eine Rolle.
Regulation
Unabhängig von Umgebungs- und Belastungsbedingungen muss die Blutversorgung zu jedem Zeitpunkt aufrechterhalten bleiben. Es muss sichergestellt werden, dass Herzaktion und Blutdruck immer bestmöglich reguliert werden, alle Organe ein Mindestmaß an Blut erhalten und der Blutstrom entsprechend den Bedürfnissen von den ruhenden hin zu den aktiven Organen verteilt wird, da eine Maximalversorgung aller Organe zur gleichen Zeit nicht möglich ist. Würden alle Organe gleichzeitig maximalversorgt werden, so würde der Blutdruck stark abfallen und zum Schock führen, weil die Gesamtblutmenge dafür nicht ausreicht.
Der Körperkreislauf besteht daher aus vielen parallel geschalteten Kreisläufen, die je nach Aktivität zu- oder abgeschaltet werden können. So wird etwa nach der Nahrungsaufnahme der Verdauungsapparat vorrangig versorgt, andere Organsysteme werden gedrosselt, Hochleistungssport ist dann nicht möglich. Die Realisierung dieser Zu- und Abschaltungen erfolgt über mehrere Wege:
- Die Gefäßweite (das Lumen) der Arterien wird durch den Spannungszustand (Tonus) der glatten Muskulatur in der Gefäßwand gesteuert. Sind die Gefäße erweitert, fließt mehr Blut in das entsprechende Gebiet.
- Arteriovenöse Anastomosen: Anastomosen sind Verbindungen zwischen kleineren Blutgefäßen, in diesem Fall zwischen Arterie und zugehöriger Vene. Diese arteriovenösen Anastomosen sind verschließbar, in diesem Fall nimmt das Blut den gewohnten Weg durch die Kapillaren. Öffnen sich diese Verbindungen, so strömt ein Großteil des Blutes aufgrund des geringeren Strömungswiderstandes von der Arterie durch die Abkürzung direkt in die Vene, das Kapillarbett bekommt also weniger Blut.
- Vorkapillare Schließmuskeln: Normale Arterien können zwar ihr Lumen verengen, aber nicht bis zu einem vollständigen Verschluss. In den kleinsten Arteriolen gibt es dagegen spezielle Bildungen der mittleren Wandschicht, die als Sphincter precapillaris bezeichnet werden. Diese können das Lumen verschließen und somit den Blutfluss im sich anschließenden Kapillarbett reduzieren.
- Sperrarterien: Sperrarterien sind Arterien, die ebenfalls ihr Lumen verschließen können. Solche Sperrarterien gibt es am Penisschwellkörper. Sie sind normalerweise geschlossen und erst ihre Öffnung löst einen Blutfluss und damit die Schwellkörperfüllung (Erektion) aus.
- Drosselvenen: Drosselvenen sind Venen, die ihr Lumen einengen können. Sie kommen vor allem in der Schleimhaut des Darmes vor. Bei einer Einengung wird der Blutabfluss aus dem Darm verlangsamt und damit die Blutmenge vergrößert und die Zeit zum Übertritt der resorbierten Nährstoffe in das Blut verlängert. Außerdem sind sie im Nebennierenmark zu finden.
Kreislaufregulatorische Einrichtungen werden durch
- lokale Steuerung,
- hormonale Signale und
- neuronale Signale beeinflusst.
Lokale Steuerung
Die lokale Steuerung oder auch Autoregulation stellt zum einen das Gleichbleiben der Organdurchblutung auch bei wechselndem Blutdruck sicher, zum anderen passt sie die Durchblutung den Stoffwechselbedingungen des Organs an (zum Beispiel steigt die Durchblutung des Magen-Darm-Traktes während der Verdauung). Dies findet auf unterschiedlichen Wegen statt.
- Beim Bayliss-Effekt findet eine Kontraktion der Gefäßmuskulatur als Antwort auf eine Gefäßweitung durch eine Blutdruckerhöhung statt. Er tritt in Gehirn, Niere und Verdauungstrakt auf, nicht aber in der Haut oder der Lunge.
- Sauerstoffmangel löst eine Gefäßweitung aus, die daraus resultierende Mehrversorgung mit Blut wirkt diesem entgegen. (In der Lunge findet das genaue Gegenteil statt, eine geringe Sauerstoffsättigung hat eine Gefäßverengung zur Folge.)
- Außerdem löst das Vorkommen gewisser Stoffe im Blut lokal eine Gefäßweitung aus. Dieser lokal-metabolische Effekt wird besonders durch eine erhöhte Konzentration von Kohlendioxid, ADP, AMP, Adenosin, Wasserstoff- und Kalium-Ionen hervorgerufen. Die daraus resultierende bessere Durchblutung begünstigt den Abtransport dieser Stoffe. Besonders wichtig ist diese Art der Steuerung im Myokard und im Gehirn.
Hormonale Steuerung
Hormone wirken entweder direkt auf die Muskulatur der Gefäßwand (z. B. Adrenalin), oder sie bewirken vor Ort die Freisetzung von gefäßaktiven Substanzen (z. B. Stickstoffmonoxid, Endothelin), die dann lokal wirksam werden.
- Stickstoffmonoxid (NO) hat eine gefäßerweiternde Wirkung. Es wird aus dem Endothel (der Gefäßwand auskleidenden Zellschicht) ausgeschüttet, wenn diese durch Acetylcholin, ATP, Endothelin-1 oder Histamin stimuliert wird.
- Endothelin-1 stimuliert zum einen die Freisetzung von NO, zum anderen wirkt es lokal direkt auf die Gefäßmuskulatur, dann aber gefäßverengend. Es wird vom Endothel nach Stimulation durch Angiotensin II und Vasopressin (Antidiuretisches Hormon) freigesetzt.
- Adrenalin wirkt je nach vor Ort überwiegenden Rezeptoren gefäßverengend (α1-Adrenorezeptoren, zum Beispiel in Haut und Niere) oder gefäßweitend (β2-Adrenorezeptoren, zum Beispiel in Skelettmuskel, Myokard und Leber). β-Rezeptoren sind empfindlicher als α-Rezeptoren, werden aber beide Rezeptoren gleichzeitig ausgelöst so dominieren die α-Rezeptoren.
- Eikosanoide haben unterschiedliche Effekte auf die Gefäße. Während Prostaglandin F2 und Thromboxane A2 und B2 gefäßverengend wirken, haben Prostaglandin-E2 und Prostacyclin gefäßerweiternde Wirkungen.
- Bradykinin, Kallidin und Histamin wirken gefäßerweiternd. Außerdem setzt die Stimulation des Endothels durch Bradykinin den EDHF (endothel-derived hyperpolarizing factor, hyperpolarisierender Faktor des Endothels) frei, der Gefäßmuskelzellen hyperpolarisiert.
- Serotonin bewirkt eine Gefäßverengung, und erhöht außerdem die Durchlässigkeit der Kapillaren.
- Angiotensin II wirkt innerhalb des Renin-Angiotensin-Aldosteron-Systems gefäßverengend, ebenso Vasopressin. Diese Gefäßverengung findet im Rahmen der Regulation des Wasserhaushalts durch die Niere statt.
Neuronale Steuerung
Die neuronale Steuerung findet hauptsächlich durch den Sympathikus statt, und setzt an den kleinen Arterien und den Arteriolen oder den Venen und deren Rückstrom zum Herzen an. Der postganglionäre Neurotransmitter ist das Noradrenalin, das an die α1-Rezeptoren anbindet, und deswegen gefäßverengend wirkt. Eine Gefäßweitung wird durch Nachlassen des Sympathikotonus erreicht. Ausgenommen hiervon ist die vom Parasympathikus innervierte Weitung der Gefäße der Speicheldrüsen und der Geschlechtsorgane (Erektion). Als Transmitter wirken NO und Bradykinin. Die Steuerung durch Sympathikus und Parasympathikus findet auf zwei Arten statt: zum einen über eine Art Bedarfsmeldung der Organe, zum anderen durch eine neuronale Mitinnervation, bei der das Gehirn neben der Aktivierung bestimmter Organe gleichzeitig deren Durchblutung steuert. Durch eine Verletzung von Nerven oder deren Fehlfunktion kann es zu einem spinalen oder neurogenen Schock kommen.
Zentrale Kreislaufsteuerung
Neben der Einflussnahme auf den Tonus der Gefäße findet auch noch eine zentrale Kreislaufsteuerung in der Medulla oblongata und dem Pons statt. Dabei werden Informationen von Kreislaufsensoren ausgewertet, die den arteriellen Blutdruck, die Pulsfrequenz, den Füllungsdruck des Niederdrucksystems und den pH-Wert, Kohlendioxid- und Sauerstoff-Partialdruck des Blutes messen.
Diese Drucksensoren befinden sich in der Wand der Aorta und der inneren Halsschlagader (Dehnungs- und Druckrezeptoren im Sinus caroticus) und im Niederdrucksystem in den Hohlvenen und den Vorhöfen (Dehnungssensoren). Diese Regulation wirkt aber nur akuten Blutdruckänderungen entgegen, wie zum Beispiel beim Aufstehen aus dem Liegen. Ist der Blutdruck jedoch immer auf einem erhöhten (oder erniedrigten) Niveau, so erfolgt eine Anpassung und der „neue“ Blutdruck wird gleich gehalten.
Die Gaspartialdrücke und der pH-Wert werden von spezialisierten Sensoren (sog. Chemorezeptoren) in Paraganglien erfasst, die ebenfalls an der Halsschlagader (Glomus caroticum), der Aorta (Paraganglion supracardiale, Syn. Glomus aorticum) und der Lungenarterie liegen.
Die Informationen dieser Sensoren werden an das Kreislaufzentrum im Nachhirn (Medulla oblongata) übermittelt.
Lymphsystem
Das Lymphsystem dient dazu, Wasser und darin gelöste Stoffe aus dem Körpergewebe wieder dem Blutkreislauf zuzuführen. In Umgebung der Kapillaren wird das Gewebe auf Grund des osmotischen Drucks von Flüssigkeit aus dem Blut durchtränkt. Am Ende kehrt diese Flüssigkeit wieder in die Blutgefäße zurück. Da dieser Prozess aber nicht hundertprozentig effektiv ist, sammeln Lymphbahnen diese Flüssigkeit, jetzt Lymphe genannt, und führen sie den Venen in der Nähe des Herzens wieder zu. Auf dem Weg dorthin fließt die Lymphe durch Lymphknoten, in denen sie gefiltert wird.
Der Blutkreislauf der Säugetiere vor der Geburt
Entwicklung beim Embryo
Der Blutkreislauf ist eines der am frühesten angelegten Organsysteme des Embryos. Die Blutgefäße entwickeln sich, vom Dottersack ausgehend, aus so genannten „Blutinseln“ im embryonalen Bindegewebe. Durch die Verschmelzung der beiden Endokardschläuche am Kopfende des Embryos, verschiedene Krümmungsvorgänge und Bildung von Scheidewänden entsteht daraus das Herz mit seinen beiden Vorhöfen und Herzkammern. Das Herz gelangt erst mit dem Längenwachstum des Embryos in seine definitive Lage in der Brusthöhle.
In der Frühphase der Entwicklung des Blutkreislaufes gibt es im vorderen Bereich des Embryos zunächst vier Aorten, zwei rückenseitige (dorsale Aorten) und zwei bauchseitige (ventrale Aorten). Die dorsalen Aorten besitzen pro embryonalen Körpersegment jeweils Abgänge nach dorsal (rückenwärts) und ventral (bauchwärts), im Bereich der Urniere auch nach lateral (seitlich), die als Segmentarterien bezeichnet werden. Die Dorso- und Ventralaorten sind im Bereich der Kiemenbogen untereinander durch die sechs Kiemenbogenarterienpaare miteinander verbunden.
Nun finden im vorderen Embryonalbereich komplexe Umbildungen statt. Die ersten fünf lateralen Segmentarterien sowie die erste, zweite und fünfte Kiemenbogenarterie verschließen sich beidseitig, die dorsalen Aorten zwischen viertem und fünftem Kiemenbogen. Der Vorderabschnitt der ventralen Aorten wird damit zur definitiven äußeren Halsschlagader (Arteria carotis externa), aus der dritten Kiemenbogenarterie und dem Vorderabschnitt der dorsalen Aorten beidseitig die innere Halsschlagader (Arteria carotis interna).
Die rechte dorsale Aorta verschließt sich hinter der sechsten lateralen Segmentarterie und wird zusammen mit der vierten rechten Kiemenbogenarterie zur späteren rechten Schlüsselbeinarterie (Arteria subclavia dextra). Die linke Schlüsselbeinarterie entsteht dagegen nur aus der sechsten lateralen Segmentarterie.
Die vierte linke Kiemenbogenarterie entwickelt zum Aortenbogen (Arcus aortae), die definitive Aorta entsteht aus dessen Fortsetzung in die linke ventrale Aorta. Der Anfangsabschnitt der rechten ventralen Aorta bildet sich zum Arm-Kopf-Stamm (Truncus brachiocephalicus) um.
Die beiden sechsten Kiemenbogenarterien wachsen in die Lungenanlage ein und werden zu den Lungenarterien (Arteriae pulmonales). Rechts verliert sie ihre Verbindung zur ventralen Aorta, aus ihrem Anfangsabschnitt entsteht der Lungenstamm (Truncus pulmonalis). Die sechste linke Kiemenbogenarterie erhält jedoch ihre Verbindung zur linken ventralen, also definitiven Aorta bei. Sie bildet damit eine Kurzschlussverbindung zwischen Lungen- und Körperkreislauf, den Ductus arteriosus (Ductus Botalli). Durch die Bildung eines spiraligen Septums (Septum aorticopulmonale) im Ursprung des unpaarigen Anfangsabschnitts der dorsalen Aorten erhält die definitive Aorta Anschluss an die linke Herzkammer, der Truncus pulmonalis an die rechte Herzkammer. Diese sehr komplizierten Umbauvorgänge des Herzens und der herznahen Arterien führen gelegentlich zu Missbildungen (z. B. Fallot-Trilogie, Fallot-Tetralogie, Fallot-Pentalogie).
Die ursprünglichen ventralen Segmentarterien der nun definitiven Aorta bilden sich bis auf drei unpaare Hauptstämme (Truncus celiacus, Arteria mesenterica superior und Arteria mesenterica inferior) in der Bauchhöhle zurück. Die lateralen Segmentarterien werden zu den Nieren- (Arteria renalis) und Keimdrüsenarterien (Arteria testicularis bzw. Arteria ovarica). Lediglich die dorsalen Segmentarterien behalten ihr ursprüngliches segmentales Abgangsverhalten und bilden die Zwischenrippenarterien (Arteriae intercostales superiores) bzw. Lendenarterien (Arteriae lumbales).
Blutkreislauf beim Fötus
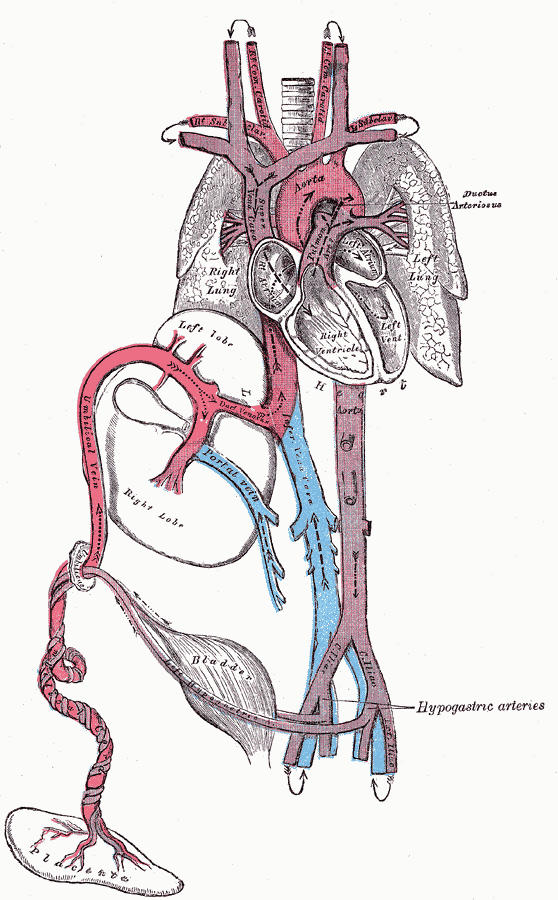
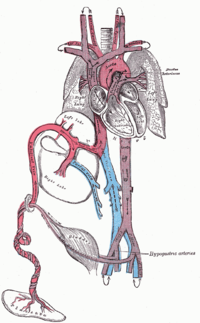 Blutkreislauf eines menschlichen Fötus
Blutkreislauf eines menschlichen FötusEtwa ab dem 21. Tag nach der Konzeption (ca. 35. Tag nach dem ersten Tag der letzten Regelblutung – gynäkologische Schwangerschaftsrechnung) beginnt das Herz des Embryos zu schlagen, in den folgenden Wochen wird auch die Lunge angelegt. Da die Lungen des Fötus im Mutterleib noch funktionslos sind, bezieht er sein sauerstoffreiches Blut über die Nabelschnur aus der Plazenta. Das sauerstoffreiche Blut gelangt aus der Nabelvene in der Nabelschnur über den Ductus venosus in die untere Hohlvene und umgeht damit zum Großteil die Leber, ein kleinerer Teil versorgt über die Pfortader die Leber mit sauerstoffreichem Blut. Dann gelangt es durch die untere Hohlvene in den rechten Vorhof. Schon in der Hohlvene mischt es sich mit dem sauerstoffarmen Blut aus dem Körperkreislauf und wird zu Mischblut. Ein Teil strömt durch das Foramen ovale in den linken Vorhof, wird in die linke Herzkammer gepumpt und verlässt das Herz durch die Aorta, um zuerst das Gehirn, das am empfindlichsten auf Sauerstoffmangel reagiert, und den oberen Teil des Körpers zu versorgen. Aus der rechten Kammer gelangt das übrige Blut in den Truncus pulmonalis, ein Teil (etwa ein Drittel) wird in die noch nicht entfalteten Lungen gepumpt. Durch die geringe Sauerstoffversorgung der Lunge sind die Lungengefäße verengt, was den Fließwiderstand erhöht. Die restlichen zwei Drittel des sauerstoffangereicherten Blutes gelangen vom Truncus pulmonaris noch vor der Lunge über den Ductus arteriosus in die Aorta (Rechts-Links-Shunt) hinter den Abgängen zum Gehirn und umgehen damit ebenfalls den Lungenkreislauf. Dieses Mischblut versorgt den unteren Teil des Körpers, bis der größte Teil über die von den inneren Beckenarterien abgehenden Nabelarterien wieder in die Plazenta fließt, wo er mit Sauerstoff angereichert wird.
Umbildungen nach der Geburt
Bei der Geburt endet die Versorgung durch die Plazenta. Dies lässt den Kohlendioxidgehalt im Blut ansteigen, was durch Chemorezeptoren einen starken Anreiz zum Atmen erzeugt. Durch das Heben des Brustkorbs sinkt der Druck innerhalb des Brustkorbes. Dies führt zum Leersaugen von Plazenta und Nabelvene und zur Entfaltung der Lungen. Da diese nun das Blut mit Sauerstoff anreichern, weiten sich die Gefäße in der Lunge, was den Gefäßwiderstand reduziert. Deshalb gelangt mehr Blut in die Lungen, die Flussrichtung im Ductus arteriosus kehrt sich um. Bis zu dessen Schließung wird die Lunge noch kurze Zeit mit Aortenblut versorgt. Nach dem Verschluss wird der Ductus arteriosus zum Ligamentum arteriosum. Während die Blutmenge im rechten Vorhof durch den Wegfall des Zuflusses aus der Plazenta abnimmt, steigt sie im linken Vorhof durch die Versorgung der Lunge. Das resultierende Druckgefälle und die Verringerung gefäßverengender Prostaglandine führen dazu, dass sich das Foramen ovale ebenfalls innerhalb der ersten zwei Wochen nach der Geburt verschließt. Ebenso verschließt sich der Ductus venosus.
Krankheiten des Kreislaufsystems
Herz-Kreislauf-Erkrankungen führen heute in den Industrienationen die Todesursachenstatistik mit Abstand an. In Deutschland wird mit leicht abfallender Tendenz knapp die Hälfte aller Todesfälle auf Krankheiten des Kreislaufsystems zurückgeführt.
Während bei Kindern und jungen Erwachsenen Herz-Kreislauf-Erkrankungen selten sind und die angeborenen Herzfehler im Vordergrund stehen, führt hauptsächlich die mit dem Alter zunehmende Arteriosklerose zu einem altersabhängigen Anstieg der Prävalenz für Herzinfarkte, Schlaganfälle und andere Durchblutungsstörungen. Unter den Herzerkrankungen (vgl. Kardiologie) sind Durchblutungsstörungen des Herzmuskels (Koronare Herzkrankheit) und Herzklappenfehler am häufigsten anzutreffen, bei den Gefäßerkrankungen (vgl. Angiologie) sind es die arterielle Verschlusskrankheit (AVK) der Arterien und das Krampfaderleiden (Varikosis) sowie die Thrombose bei den Venen. Der Bluthochdruck (arterielle Hypertonie) gehört zu den häufigsten chronischen Erkrankungen. Er ist die zweithäufigste Diagnose bei Hausärzten und gilt als bedeutsamster Risikofaktor für Herz-Kreislauf- und Nierenerkrankungen.
Forschungsgeschichte
Im 4. Jahrhundert v. Chr. wurden die Herzklappen durch einen Arzt der hippokratischen Schule entdeckt. Ihre Funktion wurde aber zu diesem Zeitpunkt noch nicht erkannt. Da sich Blut nach dem Tod in den Venen sammelt, erscheinen Arterien leer. Deswegen vermuteten antike Anatomen, dass sie mit Luft gefüllt seien und eine Rolle im Lufttransport spielen. Herophilos von Chalkedon unterschied bereits zwischen Venen und Arterien, glaubte aber, dass der Puls selbstständig durch Letztere erzeugt würde. Erasistratos beobachtete, dass am Lebenden durchtrennte Arterien bluten. Er vermutete, dass entweichende Luft durch aus kleinen Verbindungsadern zwischen Venen und Arterien nachströmendes Blut ersetzt wird. Somit war er der erste, der Kapillaren postulierte, aber mit entgegengesetztem Blutfluss. Im 2. Jahrhundert wusste Galenos (129–199) bereits, dass Blutgefäße Blut transportieren und unterschied dunkleres venöses von arteriellem Blut, welches heller und dünner ist. Beiden schrieb er verschiedene Aufgaben zu. Wachstum und Energie kämen von in der Leber aus Galle gebildetem venösen Blut, während aus dem Herz kommendes, arterielles Blut Vitalität durch enthaltene Luft brachte. Das Blut floss laut seinen Vorstellungen aus beiden Organen in alle Teile des Körpers, wo es verbraucht wurde, ohne dass ein Rückstrom zu Herz oder Leber stattfand. Das Herz selbst hat keine Pumpfunktion, sondern saugt das Blut in der Diastole ein. Der Bluttransport selbst findet durch die Pulsierungen der Arterien statt. Galen glaubte, dass arterielles Blut aus venösem Blut gebildet wird, welches durch „Poren“ in der Scheidewand zwischen den Kammern aus der rechten in die linke Herzkammer sickert.
Im 13. Jahrhundert entdeckte Ibn an-Nafis, ein arabischer Arzt und Anatom (1210/1213–1288), als Erster, dass das Blut in einem Kreislauf durch die Lunge fließt. Seine Erkenntnisse, die als Zeichnungen bis in die heutige Zeit überliefert sind, gelangten jedoch nicht bis in den europäischen Raum. 1552 beschrieb Michael Servetus (1511–1553) dasselbe Phänomen wie Ibn an-Nafis, das durch Realdo Colombo bewiesen wurde. Doch auch diese Ergebnisse wurden von der Allgemeinheit nicht anerkannt.
1628 wurde durch William Harvey (1578–1657) der Blutkreislauf erstmalig korrekt beschrieben, nachdem für 14 Jahrhunderte die Lehre Galens die medizinische Lehrmeinung bestimmt hatte. Harvey stellte seine Überlegungen auf Grund der Entdeckung der hydraulischen Funktionsweise der Venenklappen durch seinen Lehrer, den Italiener Hieronymus Fabricius Aquapendente an, da er eine Verbindung zur Funktion des Herzens suchte. Er fand sie in der Kreislauftheorie, die er 1628 veröffentlichte. Diese Arbeit begann, die Fachwelt zu überzeugen. Wie das Blut vom arteriellen in den venösen Schenkel kommt, konnte allerdings erst Marcellus Malpighi mit seiner Entdeckung der Kapillaren erklären.
Literatur
- N. A. Campbell, J. B. Reece: Biologie. Pearson Studium, München 2006 (6. Aufl.). ISBN 3-8273-7180-5
- J. R. Levick: Physiologie des Herz-Kreislauf-Systems. UTB, Stuttgart 1998. ISBN 3-8252-8129-9
- F.-V. Salomon, H. Geyer, U. Gille: Anatomie für die Tiermedizin. Enke, Stuttgart 2004. ISBN 3-8304-1007-7
- Stefan Silbernagl, Agamemnon Despopoulos: Taschenatlas der Physiologie. Thieme, Stuttgart 2003. ISBN 3-13-567706-0
Weblinks





Wikimedia Foundation.