- Evolutionsbiologie
-
Evolution ist die Veränderung der vererbbaren Merkmale einer Population von Lebewesen von Generation zu Generation. Diese Merkmale sind in Form von Genen kodiert, die bei der Fortpflanzung kopiert und an den Nachwuchs weitergegeben werden. Durch Mutationen entstehen unterschiedliche Varianten (Allele) dieser Gene, die veränderte oder neue Merkmale verursachen können. Diese Varianten sowie Rekombinationen führen zu erblich bedingten Unterschieden (Genetische Variabilität) zwischen Individuen. Evolution findet statt, wenn sich die Häufigkeit dieser Allele in einer Population (die Allelfrequenz) ändert, diese Merkmale in einer Population also seltener oder häufiger werden. Dies geschieht entweder durch Natürliche Selektion (unterschiedliche Überlebens- und Reproduktionsrate aufgrund dieser Merkmale) oder zufällig durch Gendrift.
Natürliche Selektion tritt auf, weil Individuen mit Merkmalen, die für das Überleben und die Fortpflanzung vorteilhaft sind, mehr Nachwuchs produzieren können als Individuen ohne diese Merkmale. Daher werden sie mehr Kopien ihrer vererbbaren Merkmale in die nächste Generation einbringen. Dies führt dazu, dass vorteilhafte Merkmale im Laufe der Zeit häufiger werden, während unvorteilhafte seltener werden. Durch diesen Prozess können über viele Generationen unterschiedliche Anpassungen an Umweltbedingungen entstehen. Wenn genetische Differenzen innerhalb oder zwischen Populationen von Lebewesen einer Art immer zahlreicher werden, kann sich diese Art in neue Arten aufspalten (Artbildung, Speziation). Bestimmte gemeinsame Merkmale aller Lebewesen legen nahe, dass alle bekannten Arten von einer einzigen ursprünglichen Art abstammen (genauer: von einem ursprünglichen Genpool) und durch diesen Prozess der allmählichen Verstärkung von Unterschieden entstanden sind (Deszendenz).
Die Theorie der Evolution durch Natürliche Selektion wurde erstmals ausführlich von Charles Darwin in seinem 1859 erschienenen Buch The Origin of Species dargestellt. In den 1930er Jahren wurde die von Darwin herausgestellte natürliche Selektion mit den mendelschen Regeln zur Vererbung verbunden, daraus entstand die Synthetische Theorie der Evolution. Mit ihrer außerordentlichen erklärenden und vorhersagenden Kraft wurde diese Theorie zum zentralen organisierenden Prinzip der modernen Biologie. Sie liefert die Erklärung für die Vielfalt des Lebens auf der Erde.
Der Evolutionsbiologe Ernst Mayr bezeichnete die Evolution als den wichtigsten Begriff in der gesamten Biologie.[2]
Inhaltsverzeichnis
Geschichte der Evolutionstheorie
- Hauptartikel: Evolutionstheorie, Geschichte der Evolutionstheorie
 Charles Darwin im Alter von 51 Jahren, kurz nach der Veröffentlichung des Buches The Origin of Species
Charles Darwin im Alter von 51 Jahren, kurz nach der Veröffentlichung des Buches The Origin of SpeciesIdeen zur Evolution, wie die gemeinsame Abstammung und die Umwandlung von Arten, gab es mindestens seit dem 6. Jahrhundert vor Christus, wo sie von dem griechischen Philosophen Anaximander vertreten wurden.[3] Eine größere Vielfalt solcher Ideen wurde im 18. Jahrhundert entwickelt und 1809 vertrat Lamarck die Auffassung, dass die Umwandlung von Arten durch Vererbung von Anpassungen geschieht, welche die Eltern während ihres Lebens erwerben (Lamarckismus). Diese Ideen wurden in England als eine Bedrohung der politischen und religiösen Ordnung betrachtet und vom wissenschaftlichen Establishment heftig bekämpft.
1858 präsentierten Charles Darwin und Alfred Russel Wallace gemeinsam zwei verschiedene Arbeiten zur Theorie der Evolution durch natürliche Selektion in der Linnean Society of London.[4] Diese Veröffentlichung wurde wenig beachtet, ganz im Gegensatz zu dem von Darwin 1859 veröffentlichten Buch The Origin of Species, welches die Theorie sehr ausführlich darlegt und zu einer immer größeren Akzeptanz der Erkenntnis führte, dass es Evolution wirklich gab. Darwins spezifische Thesen zur Evolution, wie der Gradualismus und die natürliche Selektion, stießen anfangs auf erhebliche Widerstände. Anhänger Lamarcks argumentierten, dass zum Beispiel Enten ihre Schwimmhäute durch ihre ständigen Versuche zu paddeln erworben hätten, und nicht durch einen Selektionsprozess, bei dem Enten mit etwas Haut zwischen den Zehen Enten ohne solche Häute im natürlichen Wettbewerb überlegen waren. Da die Experimente zur Unterstützung des Lamarckismus jedoch ohne positives Ergebnis blieben, wurde diese Theorie zugunsten der Natürlichen Selektion Darwins fallen gelassen.
Darwin konnte jedoch nicht erklären, wie Merkmale von Generation zu Generation weitergegeben werden und warum sich Variationen dieser Merkmale nicht durch Vererbung vermischten. Der Mechanismus dafür wurde 1865 von Gregor Mendel geliefert, dessen Forschungen offenlegten, dass bestimmte Eigenschaften in einer genau definierten und vorhersagbaren Weise vererbt werden.[5]
Als Mendels Arbeiten im Jahr 1900 wiederentdeckt wurden, führten unterschiedliche Meinungen über Voraussagen der ersten Genetiker und Biostatistiker zur Geschwindigkeit der Evolution zu einem tiefen Graben zwischen dem mendelschen und dem darwinschen Modell der Evolution. Dieser Graben wurde schließlich in den 1930er Jahren durch die Arbeit von Biologen wie Ronald Fisher überwunden. Das Endergebnis war eine Kombination der Darwinschen Natürlichen Selektion mit den mendelschen Regeln, die Synthetische Theorie der Evolution, die auch als Neodarwinismus bezeichnet wird.[6]
Schließlich wurde durch die Identifikation der DNA als das genetische Material durch Oswald Avery im Jahr 1944 und die Entschlüsselung der Struktur der DNA durch James Watson und Francis Crick im Jahr 1953 die physische Basis der Vererbung geklärt. Seitdem sind Genetik und Molekularbiologie zentrale Bestandteile der Evolutionsbiologie.[7] Die synthetische Evolutionstheorie definiert Evolution als die zeitliche Änderung der relativen Allelhäufigkeiten (Allelfrequenzen) in einer Population.[8]
Grundlagen
Vererbung
Hauptartikel Genetik
 Ein Teil eines DNA-Moleküls.[9]
Ein Teil eines DNA-Moleküls.[9]Gregor Mendel zeigte anhand von Erbsen, dass Vererbung in eng definierten (diskreten) Einheiten erfolgt. Er zeigte, dass Merkmale von den Eltern an die Nachkommen vererbt werden, und dass diese Merkmale diskret sind: Wenn ein Elternteil runde und der andere faltige Erbsen hatte, dann zeigte der Nachwuchs nicht ein Gemisch, sondern entweder runde oder faltige Erbsen. Mendel wies außerdem nach, dass die Merkmale der Eltern in einer genau definierten und vorhersagbaren Weise an die Nachkommen vererbt wurden, nämlich nach den mendelschen Regeln. Seine Forschungen waren die Basis für das Konzept der diskreten, erblichen Merkmale, der Gene.[10] Mendels Arbeiten beantworteten die lange offene Frage, warum Merkmalsvarianten in Populationen stabil bleiben.
Spätere Forschungen enthüllten schließlich die physische Basis der Gene und identifizierten die DNA als das genetische Material. Gene wurden neu definiert als spezifische Regionen der DNA. DNA wird von Lebewesen als Chromosomen gelagert. Ein bestimmter Ort auf einem Chromosom wird als Genlocus (oder kurz Locus) bezeichnet, die Variante eine DNA-Sequenz auf einem bestimmten Locus bezeichnet man als Allel. Die Kopie der DNA erfolgt nicht perfekt und Änderungen (Mutationen) der Gene produzieren neue Allele und beeinflussen daher die Merkmale, die von diesen Genen kontrolliert werden. Diese einfache Beziehung zwischen einem Gen und einem Merkmal liegt in vielen Fällen vor, komplexe Merkmale wie zum Beispiel die Widerstandsfähigkeit gegen Krankheiten werden jedoch von vielen zusammenwirkenden Genen kontrolliert.[11]
Genetische Variabilität
Die genetische Variabilität resultiert aus Mutationen der DNA, der Wanderung von Individuen zwischen Populationen (dem Genfluss), und der Durchmischung von Genen bei der sexuellen Fortpflanzung. Bei einigen Lebensformen, wie Bakterien und Pflanzen, wird Variabilität auch durch die Mischung des genetischen Materials zwischen Arten durch horizontalen Genfluss und Hybridisierung erzeugt.[12][13] Trotz all dieser Variabilität verursachenden Prozesse sind die meisten Bereiche der DNA einer Art (das Genom) bei allen Individuen einer Art identisch.[14] Vergleichsweise kleine Änderungen des Genotyps (des Merkmale kodierenden Teiles des Genoms), können jedoch erhebliche Auswirkungen auf den Phänotyp (die Gesamtheit der genetisch bedingten Merkmale eines Individuums; vereinfacht ausgedrückt, auf das genetisch bedingte äußere Erscheinungsbild) haben. Zum Beispiel unterscheidet sich das Genom von Schimpanse und Mensch nur zu 5 % voneinander.[15]
Der Phänotyp resultiert aus der Interaktion seiner individuellen genetischen Ausstattung, seines Genotyps, mit der Umwelt. Die Variabilität der vererbbaren Merkmale innerhalb einer Population reflektiert also die Variabilität des Genoms innerhalb dieser Population. Die Frequenz einzelner Merkmalsvarianten kann in einer Population schwanken und in Relation zu anderen Allelen des Gens größer oder kleiner werden. Alle evolutionär wirksamen Kräfte agieren, indem sie diese Änderungen der Allelfrequenzen in die eine oder andere Richtung fördern. Die Variabilität eines Merkmals verschwindet, wenn ein Allel eine feste Frequenz erreicht, wenn es also entweder aus der Population verschwindet oder wenn es alle anderen, früher vorhandenen Allele ersetzt hat.[16]
Mechanismen
Hauptartikel: Evolutionsfaktoren
Die beiden grundlegenden Mechanismen evolutionärer Änderungen, also der Änderungen der Allelhäufigkeiten (Allelfrequenzen) innerhalb einer Population, sind Natürliche Selektion und Gendrift. Natürliche Selektion ist die nichtzufällige Auswahl von Allelen, die einen Überlebens- und Reproduktionsvorteil bieten. Gendrift ist dazu im Gegensatz die zufällige Stichprobe der Gene einer Elterngeneration, die eine zufällige Änderung der Häufigkeit einiger Allele verursacht.
Natürliche Selektion
Hauptartikel: Natürliche Selektion
 Der Schwanz eines Pfaus ist ein klassisches Beispiel für Sexuelle Selektion
Der Schwanz eines Pfaus ist ein klassisches Beispiel für Sexuelle SelektionNatürliche Selektion entsteht nach Darwin aus dem unterschiedlichen Reproduktionserfolg der Individuen einer Population. Sie ergibt sich aus den folgenden Tatsachen:
- Innerhalb von Populationen und zwischen Arten gibt es eine natürliche, vererbbare Variabilität
- die Anzahl der Nachkommen der Individuen ist viel höher als die Kapazität des jeweiligen Lebensraumes, es herrscht also Konkurrenz.
- Der Überlebens- und Reproduktionserfolg der Individuen einer Population ist daher unterschiedlich
- In jeder Generation geben die erfolgreich reproduzierenden Individuen ihre vererbbaren Merkmale weiter, die nicht erfolgreichen können dies nicht.
Wenn ein Merkmal die „evolutionäre Fitness“ von Individuen erhöht, dann werden diese Individuen mit höherer Wahrscheinlichkeit überleben und reproduzieren, als andere Individuen dieser Population und daher mehr Kopien ihrer Merkmale an die nächste Generation weitergeben (survival of the fittest). Umgekehrt wird ein Fitnessverlust durch ein nachteiliges Merkmal dazu führen, das dieses Merkmal seltener wird.[17][18]
Ein Spezialfall der Natürlichen Selektion ist die Sexuelle Selektion: Die Selektion auf Merkmale, deren Präsenz direkt mit dem Kopulationserfolg durch bevorzugte Partnerwahl korreliert ist.[19] Durch Sexuelle Selektion evolvierte Merkmale sind besonders bei den Männchen von Tieren verbreitet. Obwohl diese Merkmale die Überlebenswahrscheinlichkeit einzelner Männchen reduzieren können (z. B. durch behindernde Geweihe, durch Paarungsrufe oder leuchtende Farben, die Beutegreifer anlocken, oder durch Kämpfe unter den Männchen um die Weibchen), ist der Reproduktionserfolg von jenen Männchen im Normalfall höher, die deutliche, sexuell selektierte Merkmalskombinationen zeigen.[20]
Bei mehr als 200 Vogelarten und etwa 120 Säugerarten findet man soziale Strukturen, bei denen ein Teil der Individuen zumindest zeitweise auf eine eigene Reproduktion verzichtet und stattdessen Artgenossen bei deren Reproduktion unterstützt. Dies steht in scheinbarem Widerspruch zu Darwins Thesen. Untersuchungen dieser Helfersysteme haben jedoch gezeigt, das diese Hilfe meist umso stärker erfolgt, je näher die Helfer mit dem aufzuziehenden Nachwuchs verwandt sind.[21] Da ein Teil des Genoms von Helfer und aufgezogenem Fremdnachwuchs identisch ist, erreicht der Helfer also trotz Verzicht auf eigene Reproduktion eine Weitergabe eines Teils seines Genoms. Da die Selektion hier nicht mehr auf der Ebene des Phänotyps, sondern des Genotyps ansetzt, hat Richard Dawkins für diese und ähnliche Fälle den Begriff des "Egoistischen Gens" geprägt.[22] Bei eusozialen Insekten wie beispielsweise Ameisen und Sozialen Faltenwespen verzichtet der größte Teil der Weibchen lebenslang auf eine eigene Fortpflanzung. Eusozialität ist für diese Weibchen nicht mit einer verringerten evolutionären Fitness verbunden, da sie aufgrund einer genetischen Besonderheit (Haplodiploidie) mit ihren Schwestern näher verwandt sind als mit potentiellen eigenen Nachkommen. Bei der Aufzucht von Schwestern geben sie also einen größeren Teil ihres Genoms weiter als bei der Aufzucht eigener Töchter.[23]
Gendrift
Hauptartikel: Gendrift
Gendrift ist die Änderung von Allelfrequenzen von einer Generation zur nächsten, die geschieht, weil die Allele einer Generation von Nachkommen statistisch gesehen eine Zufallsstichprobe der Allele der Elterngeneration darstellen und deren Auswahl daher auch einem Zufallsfehler unterliegt. Selbst wenn keine Selektion stattfindet, tendieren Allelfrequenzen dazu, im Verlauf der Zeit größer oder kleiner zu werden, bis sie schließlich die Werte 0 % oder 100 % erreichen. Schwankungen der Allelfrequenzen in aufeinanderfolgenden Generationen können daher durch reinen Zufall dazu führen, dass einzelne Allele aus der Population verschwinden. Zwei getrennte Populationen mit anfänglich gleichen Allelfrequenzen können daher durch zufällige Schwankungen in zwei unterschiedliche Populationen mit einem unterschiedlichen Satz von Allelen auseinanderdriften.[24] Ob Natürliche Selektion oder Gendrift den größeren Einfluss auf das Schicksal neuer Mutationen haben, hängt von der Größe der Population und der Stärke der Selektion ab.[25] Natürliche Selektion dominiert in großen Populationen, Gendrift in kleinen. Schließlich hängt die Zeit, die ein Allel benötigt, um in einer Population durch Gendrift eine feste Frequenz zu erreichen (bis also 0 % oder 100 % der Individuen der Population das Allel tragen), von der Populationsgröße ab; bei kleineren Populationen geschieht dies schneller.[26]
Daraus folgt, das eine Veränderung der Populationsgröße einen extremen Einfluss auf den Verlauf der Evolution haben kann. Wenn eine Population aufgrund äußerer Einflüsse durch einen sogenannten "Flaschenhals" geht, zeitweise also bis auf wenige Individuen zusammenschrumpft, verliert sie damit auch einen großen Teil ihrer genetischen Variabilität (Genetischer Flaschenhals). Die Population wird dadurch insgesamt gleichartiger und verliert die meisten seltenen Varianten. Solche Flaschenhälse können durch Katastrophenereignisse oder Klimaschwankungen, aber auch durch Wanderungen oder die Teilung von Populationen verursacht werden.
Indizien
Evolution ist als historisches Geschehen zwar im Nachhinein rekonstruierbar, aber aufgrund der zeitlichen Dimensionen einer experimentellen Überprüfung allenfalls in kleinen Teilschritten zugänglich. Der Ablauf muss daher für fast alle Lebensformen anhand von Indizien nachvollzogen werden.
Abgestufte Ähnlichkeiten
Darwin hat postuliert, dass alle Lebewesen auf einen gemeinsamen Vorfahren oder, moderner ausgedrückt, einen gemeinsamen Genpool zurückgehen. Belege hierfür sind einerseits Merkmale, die alle Lebewesen gemeinsam haben. Zum Beispiel benutzen alle lebenden Zellen dieselben Nukleinsäuren als genetisches Material und denselben Basissatz von Aminosäuren zur Herstellung von Proteinen.[27]
Schon die ersten Anfänge der Systematik beruhten außerdem auf der Erkenntnis, dass Merkmale von Lebewesen nicht in beliebigen Kombinationen auftreten, sondern in einem System abgestufter Ähnlichkeiten. Dabei lassen sich Merkmalsgruppen gegeneinander abgrenzen, anhand derer die rezenten Organismen in Gruppen (Taxa, Einzahl Taxon), zusammengefasst (klassifiziert) und hierarchisch geordnet werden können. Dass dies möglich ist, wurde von Darwin als starkes Indiz für eine gemeinsame Abstammung aller Lebewesen gewertet. Damit diese Gruppierungen (Systematisierungen) die tatsächlichen Verwandtschaftsverhältnisse wiedergeben (Phylogenetische Systematik), wurde als zentrales Kriterium die Homologie von Merkmalen eingeführt.
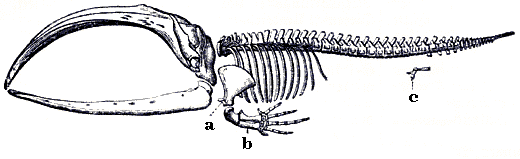 Skelett eines Bartenwals. Das c kennzeichnet die Rudimente der Hinterbeine; ein Merkmal, das die Abstammung von landlebenden Säugern belegt
Skelett eines Bartenwals. Das c kennzeichnet die Rudimente der Hinterbeine; ein Merkmal, das die Abstammung von landlebenden Säugern belegtEin Sonderfall homologer Merkmale sind morphologische Merkmale oder sogar Verhaltensweisen, die für ihre heutigen Träger keinen erkennbaren Zweck mehr erfüllen (Rudimente), beispielsweise die Reste des Hinterbeinskeletts bei Riesenschlangen und Walen. In beiden Fällen weisen diese Rudimente auf die Abstammung von vierbeinigen Tieren (Echsen bzw. Unpaarhufern) hin.
Fossile Überlieferung
Als Belege für ausgestorbene Lebewesen geben Fossilien nicht nur Auskunft über deren Morphologie, Systematik, Physiologie, Ontogenese und Ökologie. Sie bieten vor allem auch einen zeitlichen Rahmen für Evolutionsprozesse: Entsprechend der vertikalen Aufeinanderfolge fossilführender Gesteinsschichten lassen sich Gemeinschaften vorzeitlicher Lebewesen in eine zeitliche Reihenfolge bringen (Prinzip der Stratigraphie). Mit Hilfe radiometrischer Methoden kann man Gesteinen und den darin enthaltenen Fossilien auch Alter in (wenigen tausend bis vielen Millionen) Jahren zuweisen (Geochronologie).
Das Alter der Fossilien gibt Auskunft darüber, wann im Verlauf der Stammesgeschichte einzelner Gruppen sowie der Lebewesen insgesamt bestimmte Innovationen und Aufspaltungsereignisse (adaptive Radiationen) auftraten. Durch datierbare Fossilfunde ist zum Beispiel bekannt, dass (bis auf wenige Ausnahmen wie z. B. Cloudina)[28] alle Gruppen skeletttragender Tiere in einem engen Zeitfenster im frühen bis mittleren Kambrium vor etwa 540 bis 500 Mio. Jahren auftraten.[29]
Fossile Übergangsformen (Mosaikformen) (engl. "missing links") sind ein von Untersuchungen an rezenten Lebewesen unabhängiger Beweis für die Verwandtschaft zwischen systematischen Großgruppen. Berühmte Beispiele dafür sind gefiederte Dinosaurier, Archaeopteryx und die Vögel der Jehol-Gruppe als Übergangsformen zwischen Reptilien und modernen Vögeln[30] sowie Panderichthys, Tiktaalik und Ichthyostega als Übergangsformen zwischen Knochenfischen und Landwirbeltieren.[31] Im günstigsten Fall ist der Übergang zwischen der ursprünglichen und der daraus abgeleiteten Gruppe durch eine Abfolge von Fossilfunden belegt, die mit abnehmendem Alter der neuen Gruppe morphologisch immer ähnlicher werden.
Evolutionäre Trends sind in vielen Fällen innerhalb systematischer Gruppen gut dokumentiert, so z. B. bei den pferdeartigen Säugetieren: Aus vielzehigen, fuchsgroßen, laubfressenden Formen im Alttertiär sind über mehrere Zwischenstufen die heutigen Pferde hervorgegangen.
Schließlich dokumentiert die fossile Überlieferung Ab- und Zunahmen in der Diversität systematischer Gruppen. Faunenschnitte sind Massenaussterben, bei denen in geologisch gesehen kurzen Zeiträumen die Artenzahl stark reduziert wurde und manche Großgruppen völlig verschwanden oder in ihrer Vielfalt stark abnahmen. Bekanntester Faunenschnitt ist das fast vollständige Aussterben der Dinosaurier und weiterer Großgruppen am Ende der Kreidezeit mit der anschließenden Radiation der Säugetiere und der modernen Vögel.
Die folgende Zeittafel gibt einen Überblick über die wesentlichen stammesgeschichtlichen Entwicklungen der Lebewesen.
System Beginn vor Mio. Jahren Entwicklungen Quartär 1,8 Eiszeitliche Tier- und Pflanzenwelt Paläogen u. Neogen („Tertiär“) 65 Radiation der Säugetiere; erste Primaten und Hominiden Kreide 135 Entwicklung der Bedecktsamer, Massenaussterben (u. a. Dinosaurier) Jura 203 Radiation der Dinosaurier, erste Vögel (Archaeopteryx) Trias 250 Erste Säugetiere, Dinosaurier und Flugsaurier Perm 295 Radiation der Reptilien, „Säugetierähnliche Reptilien“, Massenaussterben Karbon 355 Radiation der Amphibien, erste geflügelte Insekten Devon 408 Farne, Schachtelhalme und Bärlappe entwickeln sich, Landgang der Arthropoden und Wirbeltiere Silur 435 erste Landpflanzen, Radiation der Fische Ordovizium 500 Kopffüßer erscheinen, erste „kiefertragende Fische“ Kambrium 540 „Burgess-Fauna“, Arthropoden, Chordaten (Conodonten), Wirbeltiere (kieferlose Fische) erscheinen Proterozoikum (Erdfrühzeit) 2500 Pilze, erste vielzellige Tiere: Hohltiere, Bilateria („Ediacara-Fauna“) Archaikum (Erdurzeit) 3800 Einzelliges Leben entsteht (Bakterien, Archaeen, Eukaryoten), Stromatolithen Geografische Verbreitung
Die Verbreitung von Taxa liefert in vielen Fällen Hinweise für evolutionäre Entwicklungen. Viele Taxa haben geografische Verbreitungen, die allein durch die heutigen lokalen ökologischen Verhältnisse oder Anpassungen nicht zu erklären sind. Dies gilt insbesondere für Endemiten. Ein bekanntes Beispiel für Reliktendemiten sind die Lemuren, ein Taxon innerhalb der Feuchtnasenaffen. Lemuren waren im Tertiär in Nordamerika und Eurasien weit verbreitet, sind heute aber auf Madagaskar endemisch. Sie wurden von den später evolvierten und offensichtlich konkurrenzstärkeren Trockennasenaffen (Haplorhini) überall verdrängt und konnten sich nur auf Madagaskar halten, das aufgrund einer zwischenzeitlich erfolgten geographischen Isolation von Trockennasenaffen nicht besiedelt werden konnte.
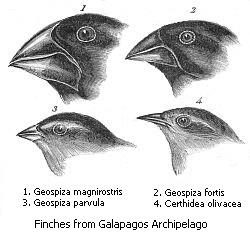
 Verschiedene Darwinfinken
Verschiedene DarwinfinkenEbenso wichtig für die Evolutionsbiologie sind Entstehungsendemiten. Berühmtestes Beispiel sind hier die Darwinfinken (Familie Emberizidae) auf den Galapagosinseln, 965 km vor der Küste Ecuadors. Auf der Inselgruppe vulkanischen Ursprungs, deren älteste Insel vor fünf bis zehn Millionen Jahren entstand, entwickelten sich aus einer vom Festland verdrifteten Vorgängerart durch adaptive Radiation 14 verschiedene Arten in drei Gattungen. Dabei änderten sich vor allem die Schnabelform, das Federkleid und die Größe der Vögel. Charles Darwin sammelte die unterschiedlichen Arten 1836 auf seiner Weltreise.
Embryonalentwicklung
Karl Ernst von Baer hat als erster erkannt, dass frühe Entwicklungsstadien verwandter Organismen einander ähnlicher sind, als die ausgewachsenen Individuen. Ernst Haeckel formulierte auf Basis dieser Erkenntnisse, dass die Ontogenese die Evolution (Phylogenese) eines Lebewesens rekapituliert (Biogenetische Grundregel). Diese Regel wird heute jedoch als zu starke Vereinfachung bezeichnet. Wiederholt werden nicht die Adultstadien, sondern die frühen Embryonalstadien der jeweiligen Vorformen einer Art. Das kann dadurch erklärt werden, dass evolutionäre Neuentwicklungen immer auf bestehende Arten und damit auf bestehende Organsysteme aufbauen. Daher schließt die Entwicklung (die Ontogenese) des Individuums einer Art auch die vorher jeweils bereits vorhandenen Schritte mit ein.
Beispiele einer solchen Wiederholung sind das Auftreten eines Kiemendarms und von Kiemenspalten bei allen Wirbeltierembryonen. Dies belegt, das alle heutigen Wirbeltiere von Fischen abstammen. Bartenwalembryonen bilden Zahnanlagen, während die adulten Individuen keine Zähne mehr haben, was darauf hinweist, dass sich die Bartenwale aus Zahnwalen entwickelt haben.
Aktuelle Forschung
- Die Koevolution
Die Entwicklung von Symbiosen ist vielfach noch nicht durch Fossilfunde abgesichert; an Modellen, wie die tiefgreifenden Abhängigkeiten von Symbiosepartnern (beispielsweise bei Flechten) entstehen konnten, besteht jedoch kein Mangel. Ebenso fehlen noch Belege für das Zustandekommen der wechselseitigen Anpassungen von Insekten und Blütenpflanzen. Überall dort, wo man aber fossile oder rezente Zwischenstufen gefunden hat, ist die Evolutionstheorie eindrucksvoll bestätigt worden.
- Die Evolution der Evolutionsmechanismen
Hier hat die Molekularbiologie in jüngerer Zeit deutlich veränderte Einsichten gebracht. Ging man in den 70er Jahren des 20. Jahrhunderts noch davon aus, dass die DNA-Sequenz direkt das entscheidende Genmaterial darstelle, so haben die Entdeckung der Introns, Exons sowie des Splicings und insbesondere des alternativen Splicings gezeigt, dass die Ursachen der genetischen Variabilität bereits auf molekularer Ebene Evolutionsprozessen unterworfen sind.
- Die Makroevolution
Die Evolution tiefgreifender Änderungen (Makroevolution), etwa auf der Ebene von Tierstämmen. Solange als Ursachen der Variabilität nur Genmutationen, Chromosomenmutationen, Genommutationen und Rekombination im Zuge der Meiose erkannt waren, war schwer vorstellbar, wie sich bestimmte Merkmale in der beobachteten Geschwindigkeit entwickelt haben könnten. Solche Erscheinungen findet man speziell bei Eukaryoten. Die Entdeckung des alternativen Splicings bei Eukaryoten hat Ende des 20. Jahrhunderts gezeigt, dass DNA-Sequenzen multifunktionell sind und – je nach Splicing – zu unterschiedlichen Proteinen führen können. Auch die Genregulation bringt neue Aspekte in die Evolutionsforschung. So kann es einen Selektionsvorteil darstellen, phylogenetisch alte und nicht zur Proteincodierung benutzte DNA-Sequenzen im Genom zu konservieren, da damit die Ausprägung neuer Merkmale durch verändertes Splicing oder Änderungen der Genregulation weitaus schneller und tiefgreifender geschehen kann, als es durch einen Austausch von DNA-Basen der Fall wäre.
Auswirkungen des Evolutionsgedankens auf Politik und Religion
Die Evolutionstheorie wurde zum Teil politisch bekämpft, zum Teil politisch missbraucht. Historisch sind vor allem die sozialdarwinistischen oder rassistischen Ausbeutungen der Theorie zu nennen (z. B. sozialdarwinistisch begründeter Rassismus in der Zeit des Nationalsozialismus). Andererseits galt Darwinismus z. B. als sozialdemokratisch (ein Vorwurf im Kaiserreich), marxistisch (bei einem Teil der NSDAP) bzw. als positiver Bezugspunkt der Arbeiterbildung oder wurde als Utopie des Züchtungsstaates mit eugenischen Theorien in Zusammenhang gebracht. Evolutionstheoretiker haben sich an den unterschiedlichsten Fronten dieses Kampfes auch politisch betätigt.
Gegner der Evolutionstheorie vertreten die Auffassung, das Entstehen des Lebens, speziell des Menschen samt seinen Eigenschaften wie dem Bewusstsein, sei extrem unwahrscheinlich und müsse deshalb durch ein intelligentes Wesen oder einen Gott eingeleitet oder vollzogen worden sein. Mit einigen religiösen Gruppierungen besteht seit der Veröffentlichung von Darwins Die Entstehung der Arten eine Kontroverse, die sich vor allem in den USA um die Vermittlung der Evolutionstheorie im schulischen Biologieunterricht dreht. Besonders im konservativen Religionsunterricht, der sich eng an den Wortlaut der biblischen Schöpfungsgeschichte hält, wird der Gegensatz zum heutigen wissenschaftlichen Kenntnisstand offensichtlich. Sofern die Kritik an Darwin und den Ergebnissen der modernen Evolutionsforschung sich auf die biblische Schöpfungsgeschichte stützt, wird sie in der Öffentlichkeit als Kreationismus bezeichnet.
Einzelnachweise
- ↑ F.D. Ciccarellli, T. Doerks, C. von Mering, C.J. Creevey, B. Snel, P. Bork: Toward automatic reconstruction of a highly resolved tree of life. In: Science. Bd. 311, Nr. 5765, 2006, S. 1283-1287. PMID 16513982
- ↑ Ernst Mayr: Das ist Evolution. C. Bertelsmann, 2003, ISBN 3-570-12013-9
- ↑ Henry Fairfield Osborn: From the Greeks to Darwin: An Outline of the Development of the Evolution Idea. Macmillan and Co.; 1905
- ↑ A. R. Wallace & C. Darwin: On the Tendency of Species to form Varieties, and on the Perpetuation of Varieties and Species by Natural Means of Selection. Jour. of the Proc. of the Linnean Society (Zoology), 3 (Juli 1858): S. 53-62 [1]
- ↑ F. Weiling: Historical study: Johann Gregor Mendel 1822-1884. Am. J. Med. Genet. 40, 1; 1991: S. 1-25; Diskussion S. 26
- ↑ Peter J. Bowler 1989: The Mendelian Revolution: The Emergence of Hereditarian Concepts in Modern Science and Society. Johns Hopkins University Press; Baltimore; 1989 ISBN 978-0801838880
- ↑ zum Beispiel U. Kutschera & K. Niklas: The modern theory of biological evolution: an expanded synthesis. Naturwissenschaften 91, Heft 6; 2004: S. 255-276.
- ↑ A. Stoltzfus: Mutationism and the dual causation of evolutionary change. Evol. Dev. 8, Heft 3; 2006: S. 304-317
- ↑ K. Edwards, D. Brown, N. Spink,J. Skelly, S. Neidle: Molecular structure of the B-DNA dodecamer d(CGCAAATTTGCG)2. An examination of propeller twist and minor-groove water structure at 2.2 A resolution. J. Mol. Biol. 226; Heft 4; 1992: 1161-1173
- ↑ H. Pearson: Genetics: what is a gene? Nature 441, Band 7092; 2006: S. 398-401
- ↑ R. Mayeux: Mapping the new frontier: complex genetic disorders. J. Clin. Invest. 115, Heft 6; 2005: S. 1404-1407
- ↑ J. Draghi & P. Turner: DNA secretion and gene-level selection in bacteria. Microbiology (Reading, Engl.) 152, Heft 9; 2006: S. 2683-2688
- ↑ J. Mallet: Hybrid speciation. Nature 446; Band 7133; 2007: S. 279-283
- ↑ R. Butlin, T. Tregenza: Levels of genetic polymorphism: marker loci versus quantitative traits. Philos. Trans. R. Soc. Lond., B, Biol. Sci. 353, Heft 1366; 1998: S. 187-98 [2]
- ↑ A. Wetterbom, M. Sevov, L. Cavelier, T. Bergström: Comparative genomic analysis of human and chimpanzee indicates a key role for indels in primate evolution. J. Mol. Evol. 63, Heft 5; 2006: S. 682-690
- ↑ W. Amos & J. Harwood: Factors affecting levels of genetic diversity in natural populations. Philos. Trans. R. Soc. Lond., B, Biol. Sci. 353: Heft 1366; 1998: S. 177-186 [3]
- ↑ R. Lande & S. J. Arnoldar: The measurement of selection on correlated characters. Evolution 37; 1983: S. 1210-1226
- ↑ J. Haldane: The theory of natural selection today. Nature 183, Heft 4663; 1959: S. 710-713
- ↑ M. Andersson & L. Simmons: Sexual selection and mate choice. Trends Ecol. Evol. (Amst.) 21; Heft 6; 2006: S. 296-302
- ↑ zum Beispiel J. Hunt, R. Brooks, M. Jennions, M. Smith, C. Bentsen, L. Bussière: High-quality male field crickets invest heavily in sexual display but die young. Nature 432, Band 7020; 2004: 1024-1027
- ↑ J. R. Krebs & N. B. Davies: Einführung in die Verhaltensökologie Blackwell; 1996: S. 343 ff.
- ↑ Richard Dawkins: The Selfish Gene 1976
- ↑ J. R. Krebs & N. B. Davies: Einführung in die Verhaltensökologie Blackwell; 1996: S. 387ff.
- ↑ R. Lande: Fisherian and Wrightian theories of speciation. Genome 31, Heft 1; 1989: S. 221-227
- ↑ M. Whitlock: Fixation probability and time in subdivided populations. Genetics 164, Heft 2; 2003: S. 767-779
- ↑ S. Otto & M. Whitlock: The probability of fixation in populations of changing size. Genetics 146, Heft 2; 1997: S. 723-733
- ↑ S. F. Mason: Origins of biomolecular handedness. Nature 311, Heft 5981; 1984: S. 19-23
- ↑ Hua, H., Z. Chen, X. Yuan, L. Zhang, & S. Xiao: Skeletogenesis and asexual reproduction in the earliest biomineralizing animal Cloudina. In: Geology, 33(4), 2005, S. 277-280.
- ↑ "kambrische Explosion": Review in Conway Morris, S.: The fossil record and the early evolution of the metazoa. In: Nature 361, Band 6409; 1993: S. 219-225.
- ↑ Review-Artikel zum Dinosaurier-Vögel-Übergang:
Norell, M. A., & X. Xu: Feathered dinosaurs. In: Annual Review of Earth and Planetary Sciences 33; 2005: S. 277-299.
Zhang, F., Z. Zhou, & G. Dyke: Feathers and 'feather-like' integumentary structures in liaoning birds and dinosaurs. In: Geological Journal 41(3-4), 2006, S. 395-404.
Zhou, Z.: The origin and early evolution of birds: Discoveries, disputes, and perspectives from fossil evidence. In: Naturwissenschaften 91(10), 2004: S. 455-471.
Zhou, Z. & F. Zhang: Mesozoic birds of china – A synoptic review. In: Frontiers of Biology in China 2(1); 2007: S. 1-14. - ↑ Long, J. A., G. C. Young, T. Holland, T. J. Senden & E. M. G. Fitzgerald: An exceptional devonian fish from australia sheds light on tetrapod origins. Nature 444, Band 7116; 2006: S. 199-202
Literatur
- Charles Darwin: Die Entstehung der Arten durch natürliche Zuchtwahl. Übersetzt von Carl W. Neumann. Nachwort von Gerhard Heberer. Reclam, Ditzingen 1986, ISBN 978-3-15-003071-4
- Charles Darwin: Die Abstammung des Menschen. Kröner, Stuttgart 2002.
- Richard Dawkins: The Selfish Gene. Reissued in new covers. Oxford University Press, Oxford 1999, ISBN 0-19-286092-5 (dt.: Das egoistische Gen)
- Richard Dawkins: The Blind Watchmaker. Reissued. Penguin, London u.a. 2000, ISBN 0-14-029122-9
- Douglas J. Futuyma: Evolutionsbiologie. Aus dem Englischen übersetzt und bearbeitet von Barbara König. Birkhäuser Verlag, Basel 1990.
- Ulrich Kutschera: Evolutionsbiologie. 3. Auflage. Verlag Eugen Ulmer, Stuttgart 2008, ISBN 3-8252-8318-6
- Ernst Mayr: Artbegriff und Evolution. Parey-Verlag, Hamburg 1967.
- Ernst Mayr: Das ist Evolution. Goldmann, München 2005, ISBN 3-442-15349-2
- Heinrich Meier (Hrsg.): Die Herausforderung der Evolutionsbiologie. 3. Auflage. Piper-Verlag, München 1992 (Serie Piper, Band 997), ISBN 3-492-10997-7
- Neil H. Shubin: The Morphogenesis and Origin of the Skeletal Pattern of the Tetrapod Limb. Harvard University, 1987
- Neil H. Shubin: Der Fisch in uns: Eine Reise durch die 3,5 Milliarden alte Geschichte unseres Körpers. Übersetzt von Sebastian Vogel. S. Fischer Verlag, Frankfurt am Main 2008, ISBN 3100720040, 9783100720047
- Volker Storch, Ulrich Welsch, Michael Wink: Evolutionsbiologie. 2. Auflage. Springer Verlag, Berlin / Heidelberg 2007, ISBN 978-3-540-36072-8
- Sven P. Thoms: Ursprung des Lebens. Fischer, Frankfurt am Main 2005, ISBN 3-5961-6128-2
- Alfred Russel Wallace: On the tendency of varieties to depart indefinitely from the original type. 1858.
- Carl Zimmer: Evolution: The Triumph of an Idea. 2001, ISBN 0-06-019906-7
Weblinks
- Evolutionsbiologie in Deutschland, umfassende Seite mit Literaturempfehlungen, Texten und weiteren Links
- Evolutionsfaktoren und Indizien, Linkliste zu speziellen Themen zum Anklicken
- ARD-Mediathek Audiobeiträge zum Thema Evolution
- Evolution - Zufall und Zwangsläufigkeit der Schöpfung Audiomitschnitte der öffentlichen Ringvorlesung an der Universität Göttingen im Wintersemester 2007/08
- Teaching about Evolution and the Nature of Science
- Henry Gee, Rory Howlett and Philip Campbell - 15 evolutionary gems Artikel über 15 Fälle der aktuellen Forschung, die die Darwin'sche Evolutionstheorie belegen. nature, 2009. doi:10.1038/nature07740


Wikimedia Foundation.