- Nematocyste
-
Die Nesselzellen, auch als Nematocyten oder Cnidozyten bezeichnet, sind ein spezieller Zelltyp, der ausschließlich bei den Nesseltieren (Cnidaria) zu finden ist. Es handelt sich dabei um Zellen, die in der äußeren Schicht (Epidermis) der Tiere eingebettet sind und zum Beutefang oder zur Abwehr von Feinden oder Konkurrenten eingesetzt werden können. Bei Reizung wird ein Nesselschlauch ausgeschleudert, der häufig ein hochwirksames Gift in das Opfer injiziert. Obwohl sie bei Menschen in der Regel nur Reizungen der Haut und leichtere Verbrennungen auslösen, sind die Nesselgifte einiger Arten so wirksam, dass sie zum Zusammenbruch des Herz-Kreislauf-Systems und somit zum Tode führen können.
Inhaltsverzeichnis
Lage der Nesselzellen
 Nesselzellen sitzen an den feinen Tentakeln dieser Schirmqualle
Nesselzellen sitzen an den feinen Tentakeln dieser SchirmqualleDie Anordnung der Zellen auf dem Körper der Tiere ist nicht gleichmäßig. In der Außenhaut, der Epidermis, finden sich Nesselzellen auf allen Strukturen, die zum Beutefang oder zur Verteidigung eingesetzt werden, in besonders hoher Dichte - man nennt diese Ansammlungen dann auch Batterien. Sie sitzen in erster Linie auf den Tentakeln, aber auch auf speziellen Strukturen: Dazu zählen etwa die Acontia der Seeanemonen (Actinaria), die normalerweise als feine Fäden im Körperhohlraum, dem Coelenteron, liegen, aber bei Gefahr durch kleine Öffnungen desselben, die Cincliden, nach außen geschleudert werden. Seeanemonen besitzen darüber hinaus zur Verteidigung gegen genetisch fremde Artgenossen Acrorhagi, fadenförmige Strukturen, die unterhalb der echten Tentakel liegen und als Waffe eingesetzt werden - nach einem Acrorhagi-Duell zieht sich die unterlegene Anemone meist mit erheblichen Gewebeschäden zurück. Steinkorallen (Scleractinia) haben zwar weder Acontia noch Acrorhagi, können aber Nesselzellen tragende Bänder, die auf den Scheidewänden des Coelenterons sitzen, nach außen ausstülpen und zum Beutefang und zur externen Verdauung einsetzen. Bei den Hydrozoen (Hydrozoa) gibt es spezialisierte Polypen ohne Mund oder Tentakel, die aber dicht mit Nesselzellen besetzt sind und nur der Verteidigung dienen. Sie heißen je nach Taxon Dactylozooide oder Nematophoren.
Lokalisiert kommen Nesselzellen bei allen Nesseltieren auch in der Innenhaut, der Gastrodermis vor. Sie dienen dort dazu, verschlungene, aber noch nicht gänzlich abgetötete Beute während des Verdauungsprozesses in gelähmtem Zustand zu erhalten.
Manche Nesseltiere verfügen über bis zu sechs Millionen Nesselzellen, die in über 1000 Batterien angeordnet sind.
Aufbau der Nesselzellen
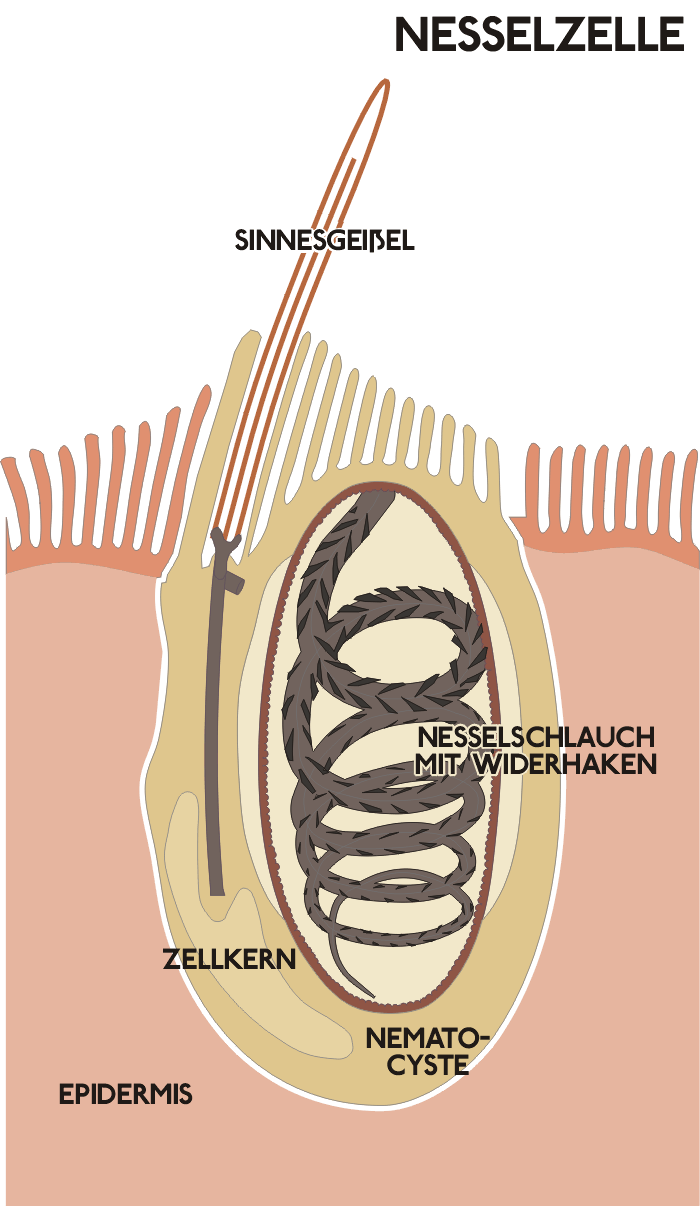
 Ursprüngliche Nesselzelle im unentladenen Zustand
Ursprüngliche Nesselzelle im unentladenen ZustandNesselzellen enthalten als Hauptbestandteil eine Nesselkapsel oder Nematocyste, die fast den gesamten Zellraum einnimmt. Sie ist von einer Kapselhülle umgeben, die durch eine zusätzliche Kollagenschicht versteift ist. Der Zellkern sowie andere Zellkompartimente liegen am Rand der Zelle zwischen dieser Kapsel und der Zellmembran. Die Kapsel selbst enthält einen mehr oder weniger aufgewickelten und 5 bis 100 Mikrometer langen Nesselschlauch, der als Einbuchtung (Invagination) der Kapselmembran anzusehen ist und abhängig vom Typ der Zelle mit unterschiedlichen Strukturen wie Stiletten, Stacheln oder Klebeelementen ausgestattet ist. Die Nesselzelle ist flüssigkeitsgefüllt und enthält einzelne Aminosäuren, einige Proteine, oft mit toxischer Wirkung, sowie in hoher Konzentration saure Peptide wie Poly-Gamma-Glutaminsäure.
Am oberen, der Außenwelt zugerichteten, Ende besitzt die Zelle einen Rezeptorpol, der auf mechanische Reize reagiert (Mechanorezeptor). Bei den Blumentieren (Anthozoa) findet man als Rezeptor den wahrscheinlich ursprünglicheren Typ. Er besteht aus einer normalen Sinnesgeißel, der Cilie, mit zugehörigem Basalkörper und Cilienwurzel sowie einem akzessorischen Zentriol. Dieser Grundaufbau entspricht dem einer Reihe anderer Mechanorezeptoren wie etwa den Hautsensoren verschiedenster Tiergruppen oder auch den Rezeptoren im Innenohr der Wirbeltiere. Die anderen Taxa der Nesseltiere besitzen demgegenüber ein so genanntes Cnidocil, welches aus einer speziell umgewandelten, versteiften Cilie mit darum angeordneten Mikrovilli (Stereocilien) besteht. Einen Basalkörper besitzt auch diese Struktur, die Wurzel und das akzessorische Centriol fehlen.
Die Nesselzelle ist eingebettet in die äußere Zellschicht der Nesseltiere, die Epidermis, wobei immer mehrere Nesselzellen mit einer Epithelmuskelzelle verbunden sind. An den Kontaktstellen verlaufen in beiden Zellen vertikal zu der Zelloberfläche feine, aus Proteinen bestehende Röhrchen, die Mikrotubuli, die offensichtlich eine mechanische Verbindung der beiden Zelltypen darstellen. In diesem Zellkomplex enden außerdem Nervenzellen, deren Enden Vesikel mit chemischen Neurotransmittern enthalten können. Sowohl der Epithelmuskelzelle als auch der Nervenendigung kommt wahrscheinlich eine Funktion bei der Aktivierung der Nesselzelle zu: Lange Zeit wurde angenommen, Nesselzellen seien autonome Systeme, die auf einen entsprechenden Reiz aus der Umgebung unabhängig von allen anderen Zellen die Entladung auslösen. Aus diesem Grunde wurden sie auch als unabhängige Rezeptor-Effektoren bezeichnet, um darauf hinzuweisen, dass sie sowohl Reize aus der Umgebung empfangen (Rezeptor-Funktion) als auch nachfolgend eine entsprechende Aktion auslösen (Effektor-Funktion). Dagegen gilt es heute als sehr wahrscheinlich, dass die Auslösung der Zelle durch umfangreichere Rezeptor-Effektor-Komplexe gesteuert wird, in welche die individuelle Nesselzelle zwar eingebettet ist, zu der aber auch noch weitere Hilfszellen und eventuell sensorische Nervenzellen gehören.
Funktion der Nesselzellen
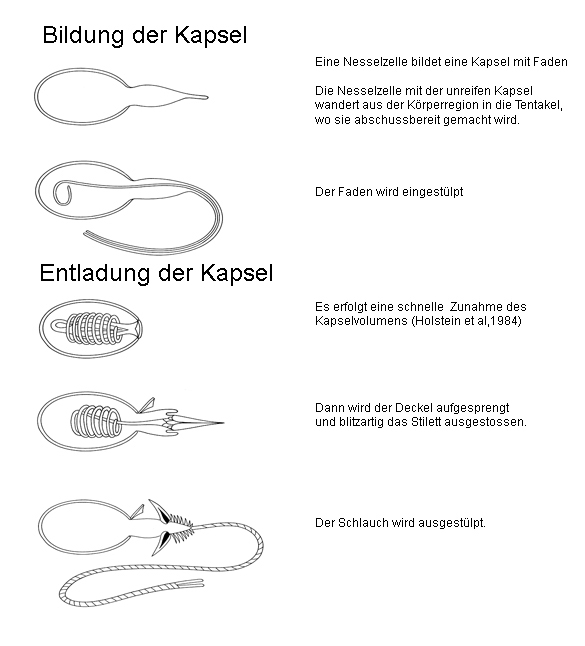
 Bildung und Entladung der Nesselkapsel
Bildung und Entladung der NesselkapselDie Funktion der Nesselkapsel ist es, den darin enthaltenen Nesselschlauch auszuschleudern. Einige Kapseltypen (z. B. Volventen) bilden einen klebrigen Wickelschlauch, der relativ langsam ausgestoßen wird. Andere Kapseltypen (z. B. Penetranten) bilden zusätzlich zum Nesselschlauch ein Stilett aus, das extrem schnell ausgestoßen wird und ein Loch in die Körperwand der Beute schlägt. Durch dieses Loch wird dann der Nesselschlauch ausgestülpt und das Nesselgift in die Beute injiziert (Abb.). Der Nesselschlauch mit den zelltypischen Stiletten, Stacheln oder Klebeelementen wird mit einer Beschleunigung von (nach neuesten Messungen von Mai 2006) zu Beginn 1,4 Millionen g (Nesselzelle einer Hydra) ausgeschleudert. Die Beschleunigung steigert sich dann innerhalb von 700 Nanosekunden auf 5,41 Millionen g. Die Spitze erreicht eine Endgeschwindigkeit von 9,3 bis 18,6 Metern pro Sekunde. Der ganze Vorgang dauert je nach Zelltyp weniger als zwei Millisekunden oder einige Sekunden. Bisher war man der Meinung, dass ein hoher osmotischer Innendruck von ca. 140 bar die schnelle Entladung und die hohe Durchschlagskraft ermöglicht. Der Druck soll durch die in der Kapsel (der Penetrante von Hydra) nachgewiesene Poly-Gamma-Glutaminsäure (ca. 20 Untereinheiten pro Molekül) erzeugt werden. Der kolloidosmotische Druck wurde jedoch falsch berechnet, denn jedes Molekül Poly-Gamma-Glutaminsäure trägt nicht 20fach sondern nur einfach zum osmotischen Druck bei. Der vorhandene Druck ist demnach mindestens um den Faktor 20 geringer als bisher angenommen. Neue Messungen ergaben im Inneren der Kapsel einen sehr niedrigen pH-Wert. Bei diesem pH-Wert ist die Poly-Gamma-Glutaminsäure nicht dissoziiert, sie bildet über Wasserstoffbrücken Aggregate und trägt daher kaum zum osmotische Wert bei. Wahrscheinlich gibt es in der schussbereiten Kapsel gar keinen Überdruck.
 Coulomb Explosion
Coulomb ExplosionNach neuerer Vorstellung wird die extrem schnelle Entladung z. B. der Stenothelen durch eine Coulombexplosion hervorgerufen. In einer schussbereiten Nesselkapsel herrscht ein niedriger pH-Wert und daher sind die Poly-Glutaminsäure-Moleküle entladen und dicht gepackt. Das Auslösen der Entladung lässt den Protonengradienten zum Zytoplasma der Nesselzelle zusammenbrechen. Protonen verlassen daraufhin die Kapsel. Dadurch sind die Säuregruppen der Poly-Glutaminsäuren gleichzeitig und schlagartig geladen, sie stoßen sich ab: eine sog. Coulombexplosion findet statt. Der Prozess ist deshalb so extrem schnell, weil die Protonen sich im Wasser viel schneller bewegen als andere Ionen (es wird nur die Ladung transportiert) und weil ein Protonen-Konzentrationsgefälle von mehreren Zehnerpotenzen zwischen dem Zytoplasma der Nesselzelle und dem Inneren der Kapsel existiert. Die Coulombexplosion führt zum Aufsprengen des Kapseldeckels und zum schnellen Ausstoßen des Stiletts. In einer zweiten, längeren Phase strömen aufgrund der jetzt vorhandenen negativen Ladungen viele Ionen und Wasser in die Kapsel und bewirken einen weiteren, relativ langsamen Druckanstieg durch osmotische Kräfte. Jetzt wird der Nesselschlauch aus- und umgestülpt. In einer dritten, möglicherweise mit der zweiten gleichzeitig stattfindenden Phase, könnten durch den pH-Wert Anstieg die Proteine der Schlauchwand ihre Konformation so ändern, dass sie die Ausstülpung des Schlauches unterstützen. Der hohe Innendruck ist also nicht permanent vorhanden, sondern wird erst im Moment des Auslösens erzeugt.
Kontrolle der Nesselzellen
Sobald die Nesselkapsel entladen ist, degeneriert die Nesselzelle, stirbt ab und wird innerhalb von 48 Stunden ersetzt. Der Hydrozoen-Polyp Hydra littoralis verliert auf diese Weise beim Fang eines Salinenkrebses (Artemia salina) etwa ein Viertel seiner auf den Tentakeln sitzenden Nesselzellen. Um unnötige Nesselzellverluste, etwa bei der Berührung durch harmlose Gegenstände zu vermeiden, besitzen Nesseltiere daher verschiedene Anpassungen. Zum einen sind meist zwei voneinander unabhängige Reizungen, zum Beispiel mechanischer und chemischer Art, notwendig, um ein großflächiges Entladen von Nesselzellen auszulösen. Eine geringe Zahl "feuert" allerdings schon bei kleinsten Berührungen, während chemische Reizung allein grundsätzlich nicht zur Auslösung führt. Die mechanischen Rezeptoren reagieren darüber hinaus auf Schwingungsfrequenzen und -amplituden besonders empfindlich, mit der sich die Gliedmaßen ihrer Beutetiere bewegen. Die Gegenwart spezieller Aminosäuren und Zucker oder kleiner Peptide wie Glutathion, die auf schleimige Absonderungen von Beute oder Fressfeinden hinweisen könnten, setzt die Auslöseschwelle ebenfalls herab.
Es gibt Hinweise darauf, dass beide Informationen integriert werden: Ein für bestimmte Beutetiere charakteristischer chemischer Reiz stellt also die mechanischen Rezeptoren auf die Wahrnehmung einer dazu passenden Schwingungsfrequenz ein. So liegt die normale Auslösefrequenz bei Süßwasserpolypen der Gattung Hydra zwischen 50 und 75 Hertz, in zuckerhaltiger Lösung dagegen bei nur 5 bis 30 Hertz, in Gegenwart großer Mengen der Aminosäure Prolin wiederum bei 90 bis 100 Hertz. Diese Daten werden etwa wie folgt gedeutet:
Kleinere Beutetiere wie etwa Krebstiere aus dem Plankton mit ihren schleimigen Absonderungen geben immer auch kleinere Mengen an Zuckern an ihre Umgebung ab. Diese setzen zunächst die Reizschwelle der Nesselzellen herab, so dass es bei mechanischer Berührung zu einem Entladen zahlreicher Zellen in das Beutetier kommt. Verwundet setzt dieses aus der Wunde zusätzliche Zucker und kleinere Peptide frei und beginnt gleichzeitig mit heftigen Strampelbewegungen, die ihre höchste Amplitude im niederfrequenten Bereich bei weniger als 30 Hertz aufweisen. Da dies dem Auslöseprofil in der Gegenwart von Zuckern entspricht, entladen sich daraufhin weitere Nesselzellbatterien in die Beute. Diese ist nun stark verwundet; aus der Körperhöhlung treten verstärkt Aminosäuren wie Prolin aus. Dies wiederum erhöht die Auslösefrequenz der Nesselzellen nun auf so hohe Werte, dass sie außerhalb des Bewegungsspektrums der Beute liegt - mit der Folge dass keine weiteren Nesselzellen mehr entladen werden. Der Einsatz von Nesselzellen ist also ökonomisch auf frische und verwundete, aber noch kämpfende Beutetiere beschränkt.
Nicht zuletzt hängt die Empfindlichkeit der Nesselzellen auch vom Ernährungszustand ihrer Träger ab; es ist bekannt, dass die Auslöseschwelle bei ausgehungerten Tieren erheblich reduziert ist; diese Information wird vermutlich durch das Nervensystem vermittelt.
Bildung von Nesselzellen
Die Nesselzellen bilden sich aus den interstitiellen Zellen (I-Zellen) an der Basis der Epidermis. Dabei handelt es sich um embryonal angelegte Zellen, die sich in alle Zelltypen der Nesseltiere mit Ausnahme der Keimzellen und der Epithelzellen entwickeln können. Die I-Zellen teilen sich, und die Bildung der Nesselzellen, deren Bildungszellen dann als Cnido- oder Nematoblasten bezeichnet werden, beginnt mit einer räumlichen Vergrößerung des Golgi-Apparates und des rauen endoplasmatischen Reticulums in der interstitiellen Zelle. In diesen Zellkompartimenten wird das Material für die spätere Nesselkapsel gebildet, die zunächst als homogene Matrix erscheint und durch Verschmelzung mit Vesikeln des Golgi-Apparates an Größe zunimmt. Dabei handelt es sich also strenggenommen um ein Sekretionsprodukt der sich bildenden Nesselzelle. Offensichtlich als Transportstrukturen für die Vesikel bilden sich [Mikrotubulus|Microtubuli] zwischen dem Golgi-Apparat und der Matrix.
In der Matrix bildet sich ein randlicher Bezirk aus, der die spätere Kapsel bildet, sowie ein zentraler Bereich, in dem sich (abhängig vom Nesselzelltyp) Stilette, oder andere Strukturen herausbilden. Auch der Nesselfaden bildet sich im Inneren der Zelle. Am oberen Rand der Kapsel setzt sich ein Deckel (Operculum) gegenüber der Restkapsel ab. Zum Abschluss der Nesselzellenbildung wird die Größe des Golgi-Apparates wieder reduziert und das Endoplasmatische Reticulum zerfällt in freie Ribosomen und einzelne Vesikel. Die Zelle wandert nun an ihre endgültige Position innerhalb der Epidermis und der Binnendruck der Kapsel wird aufgebaut. Die Bildung dieser Zellen ist irreversibel.
Typen von Nesselzellen
Insgesamt sind bis heute etwa 27 verschiedene morphologische Typen von Nesselzellen bekannt, die bei unterschiedlichen Vertretern der Nesseltiere gefunden wurden und somit für die systematische Einordnung der Tiere eine große Rolle spielen. Diese lassen sich wiederum in etwa 50 bis 60 Kapselformen unterteilen. Dabei kommen bei einer Art in der Regel mehrere Typen gleichzeitig vor. So besitzen etwa die Süßwasserpolypen der Gattung Hydra vier verschiedene Nesselzelltypen. Die Gesamtheit der Nesselzellen einer Art wird als Cnidom bezeichnet.
Unterschieden werden zunächst drei verschiedene Grundtypen von Nesselkapseln, die als Nematocysten im engeren Sinne, Spirocysten und Ptychocysten bezeichnet werden.
Am besten erforscht sind bisher die Nematocysten; hier lassen sich zwei Gruppen aufgrund des Aufbaus des Nesselschlauches unterscheiden:
- Die Astomocniden besitzen am Ende des Nesselschlauches keine Öffnung. In diese Gruppe gehören die Desmonemen oder Volventen (Wickelkapseln), die einen langen, in der Kapsel aufgewickelten Faden besitzen, mit dem Beutetiere oder Teile derselben umwickelt und festgehalten werden. Auch die Rhopalonemen mit einem keulig endenden Schlauch werden dieser Gruppe zugeordnet.
- Bei den Stomocniden ist immer eine Öffnung am Ende des Nesselfadens vorhanden. In diese Gruppe gehören die Haplonemen mit einem gleichmäßig aufgebauten Schlauch inklusive der Glutinanten (Klebkapseln), die etwa bei Hydra als Haftkapseln auch die Fortbewegung unterstützen. In ihrer Wirkungsweise haben diese Kapseln große Ähnlichkeit mit den Collocyten der Rippenquallen (Ctenophora), die jedoch in ihrer Entstehung und in ihrem Aufbau nichts mit den Nesselzellen gemein haben. Die größte Gruppe der Stomocniden bilden die Heteronemen, deren Nesselschlauch sich in einen Schaft und einen Faden unterteilen lässt. Dabei werden wiederum verschiedene Untertypen unterschieden wie etwa
- die Rhabdoide mit einem isodiametrischen Schaft,
- die Eurytelen mit einem keulenförmig erweiterten Schaft,
- die Birhopalide, bei denen der Schaft hantelförmig ist und schließlich
- die komplex aufgebauten Stenotelen, auch Penetranten oder Durchschlagskapseln, mit einem aus drei Dornen aufgebauten Stilettapparat. Sie enthalten in der Kapsel außerdem hochwirksame Nesselgifte, mit denen die Tiere ihre Opfer lähmen oder töten können.
Der zweite Grundtyp von Nesselkapseln sind die Spirocysten, die sich nur bei den Blumentieren (Anthozoa) finden. Sie besitzen im Gegensatz zu den Nematocysten nur eine dünne Kapselwand. Der Nesselschlauch ist wie eine Spiralfeder aufgerollt und nicht mit Dornen besetzt, sondern trägt stattdessen seitlich feine Klebefädchen. Cilien oder Cnidocile als Auslösemechanismus kommen bei den Spirocysten nicht vor.
Ptychocysten schließlich, die man nur bei Zylinderrosen (Ceriantharia) findet, sind stark abgewandelte Nesselkapseln, die nicht der Verteidigung dienen. Der klebrige Nesselschlauch ist hier nicht spiralig, sondern zickzackförmig zusammengefaltet, trägt aber anders als bei den Spirocysten keine Seitenfäden. Durch Cilien ausgelöst verfilzen sich die klebrigen Fasern und bilden so eine feste, feingewebte Hülle um den Polypen, die als Schutz und Wohnröhre dient.
Nesselgifte
Bei den Nesselgiften der Nesseltiere handelt es sich zum einen um Gifte, die auf das Nervensystem wirken, also den Neurotoxinen zugeordnet werden. Sie führen zu einer Blockade des Natrium-Ionen-Transportes an der Zellmembran der Nerven und verhindern damit die Bildung von Aktionspotenzialen. Das Resultat sind Lähmungen.
In Herzmuskelzellen kann ein anderer Effekt entstehen. Hier werden Kalzium-Ionen freigesetzt, wobei Krämpfe bis hin zum Herzversagen sowie ein Ausfall des gesamten Herz-Kreislaufsystems die Folge sein können. Gifte, die auf diese Weise wirken, bezeichnet man als Cardiotoxine.
Neben den Neurotoxinen finden sich im Nesselgift auch Gifte, die eine abbauende Wirkung auf Proteine oder eine blutzersetzende Wirkung haben und als Enzyme bereits die externe Verdauung in der Beute einleiten.
Nesselzellen und der Mensch
 Die Portugiesische Galeere kann auch Menschen gefährlich werden
Die Portugiesische Galeere kann auch Menschen gefährlich werdenDie meisten Nesselgifte rufen beim Menschen Hautreizungen oder leichtere Verbrennungen hervor, ernsthaftere Verbrennungen, Nekrosen und tiefe Wunden auf der Haut kommen relativ selten vor. Der Kontakt mit den Nesselzellen einiger Arten wie der Portugiesischen Galeere (Physalia physalis) oder der zu den allgemein hochgefährlichen Würfelquallen zählenden Seewespen (Chironex fleckeri und Chiropsalmus quadrigatus) kann jedoch tödlich verlaufen; ihr Gift ist wirksamer als Kobratoxin. So sterben in Australien jedes Jahr mehr Menschen durch Seewespen als durch Hai-Angriffe. Der Tod tritt meist innerhalb von Minuten ein und wird durch die Einwirkung der Toxine auf das Herz-Kreislauf-System herbeigeführt. Inwiefern sich die Wirkung für die Herstellung von Medikamenten zur Steigerung der Pumpkraft bei Patienten mit Herzinsuffizienz nutzen lässt, wird noch erforscht.
Bei Vergiftungen durch Nesselgifte der Seewespen werden Spülungen mit Essig oder Soda empfohlen, da Nesselzellen sowohl in saurem als auch in basischem Milieu kaum auslösen. In Notsituationen kann auch der saure pH-Wert menschlichen Urins genutzt werden. Auf eine Spülung mit Cola und anderen Limonaden sollte trotz des niedrigen pH-Wertes dennoch verzichtet werden, da Nesselzellen in zuckerhaltiger Lösung häufiger auslösen.
Kleptocniden
Obwohl die Nesseltiere die einzigen Tiere sind, die selbst Nesselzellen ausbilden, kann man sie auch bei einigen anderen Meerestieren finden. Dies ist dann der Fall, wenn sich die Tiere von den Polypen oder Medusen der Nesseltiere ernähren und Mechanismen entwickelt haben, die Nesselzellen aufzunehmen. Nesselzellen werden dann nicht verdaut, sondern in die eigene Epidermis eingelagert und dienen dort nun dem Schutz der Tiere vor Fressfeinden oder dem Beutefang. Nesselzellen, die auf diese Weise bei Nicht-Nesseltieren vorkommen, werden als Kleptocniden, also "geklaute Nesselzellen", bezeichnet.
Eine neue Vorstellung (s. oben Abschnitt 'Funktion der Nesselzellen') liefert eine Erklärung: Für die Bildung und die Funktion der Nesselkapsel ist eine Ansäuerung der Kapselmatrix von zentraler Bedeutung. Nacktschnecken, Plattwürmer und andere Organismen, die Nesseltiere fressen, versuchen auch die (unfertigen) Nesselkapseln zu verdauen. Dabei werden die Kapseln angesäuert und werden so -unfreiwillig- abschussbereit gemacht. Wie diese Kapseln an den Abschussort gelangen, ist ungeklärt.
So kommen Nesselzellen etwa bei verschiedenen Rippenquallen (Ctenophora) vor, was die Annahme einer nahen Verwandtschaft dieser beiden Gruppen lange Zeit unterstützte. Auch verschiedene Meeresschnecken und Plattwürmer besitzen Nesselzellen, die in die Epidermis eingelagert sind.
Stammesgeschichtlicher Ursprung der Nesselzellen
Die Herkunft der Nesselzellen ist bis heute unbekannt. Vorgeschlagen wurde ein symbiogenetischer Ursprung, also eine symbiotische Beziehung zweier Partner, bei der beide schließlich ihre Eigenidentität verloren haben.
Demnach wäre ein einfach gebautes Gewebetier, etwa nach Art von Trichoplax adhaerens eine Symbiose mit einem Protisten-Partner, etwa aus den Reihen der (heute) parasitischen Sporozoen oder der Microsporidia eingegangen, der bereits über eine cnidenähnliche Struktur verfügte. Solange dies die evolutionäre Fitness beider Partner erhöht hätte, könnte sich durch Koevolution eine immer engere Beziehung ausgebildet haben, die schließlich zur Verschmelzung zweier verschiedener Stammlinien und zur Integration der genetischen Information in einem Zellkern geführt hätte.
Dieses Szenario ist aber hochgradig spekulativ. Zu seiner Begründung werden die folgenden Beobachtungen angeführt:
- Zwischen den gewöhnlichen Epithel-Zellen und den interstitiellen Zellen (I-Zellen), aus denen die Nesselzellen hervorgehen, bestehen große Unterschiede in Hinblick auf Morphologie, Zelldifferenzierung und Zellteilung, so dass von zwei Zellpopulationen gesprochen werden kann, die sich einen gemeinsamen Körper teilen.
- Die Zellpopulationen können getrennt voneinander existieren: Während I-Zellen im Reagenzglas unter bestimmten Bedingungen außerhalb ihrer normalen zellulären Umgebung existieren können, sind Hydra-Polypen wie auch ihre Planula-Larven, denen man sämtliche I-Zellen und deren Differenzierungsprodukte wie Nesselzellen entnommen hat, lebensfähig, solange sie mit Nährstoffen versorgt werden. Auch Zellteilungen finden weiterhin statt.
- Bei der Rekombination beider Zelltypen durch "Impfung" der auf epitheliale Zellen reduzierten Tiere mit I-Zellen bildet sich wieder ein intakter Polyp heraus.
- Die Vorgängerzellen der Nesselzellen, die Cnidoblasten, zeigen eine Form der Zellteilung, die der Merogonie parasitischer Protisten ähnelt.
- Zahlreiche Symbiosen von Nesseltieren mit Protisten, in erster Linie diversen Algen, sind bekannt.
Dennoch ist das vorstehende Modell lediglich als ein mögliches Szenario zu verstehen - die tatsächliche Entstehung der Nesselzellen muss bis auf weiteres als ungeklärt gelten.
Eine neue Vorstellung (s. oben Abschnitt'Funktion der Nesselzellen') liefert auch Erklärungen für die Entstehung der Nesselkapseln. Nach dem neuen Modell ist für die Bildung und die Funktion der Kapsel eine Ansäuerung der Kapselmatrix von zentraler Bedeutung. Vermutet wird daher, dass die Nesselkapseln ihren Ursprung in Lysosomen oder ähnlichen Zellorganellen haben. Allerdings muss in der Kapsel ein pH-Wert erreicht werden, der niedriger ist als der in Lysosomen.
Literatur
- Berking S, Herrmann K (2006) Formation and discharge of nematocysts is controlled by a proton gradient across the cyst membrane, Helgol. Mar. Res. 60, 180-188
- Hessinger DA, Lenhoff HM (Hrsg., 1988): The Biology of Nematocysts, Academic Press, San Diego
- darin speziell: Holstein T, Hausmann K: The Cnidocil Apparatus of hydrozoans: A progenitor of higher metacoans mechanoreceptors?
- Holstein T (1981): The morphogenesis of nematocysts in Hydra and Forskalia: An ultrastructural study. J. Ultrastruct. Res. 75, 276-290
- Holstein T, Tardent P (1984): An ultrahigh-speed analysis of exocytosis: Nematocyst discharge, Science 223, 830-833
- Nüchter et al. (2006): Nanosecond-scale kinetics of nematocyst discharge, Current Biology 16, R316-R318
- Schäfer W (1996): Cnidarier, Nesseltiere in Westheide W, Rieger R: Spezielle Zoologie - Teil 1: Einzeller und Wirbellose Tiere, Gustav Fischer Verlag Stuttgart, Jena
- Shostak, S (1993): A symbiogenetic theory for the origins of cnidocysts in Cnidaria, Biosystems 29, 49-58
- Shostak, S, Kollavi V: Symbiogenetic origins of cnidarian cnidocysts, Symbiosis 19, 1-29
- Watson GM, Hessinger DA (1989): Cnidocyte mechanoreceptors are tuned to the movements of swimming prey by chemoreceptors, Science 243, 1589-1591
- Watson GM, Mire-Thibodeaux P (1994): The cell biology of nematocysts, International Review of Cytology 156, 275-300
- Welsch U, Storch V (1973): Einführung in Cytologie und Histologie der Tiere, Gustav Fischer Verlag, Stuttgart



Wikimedia Foundation.