- Adaptive Radiation
-
Adaptive Radiation am Beispiel der Darwinfinken
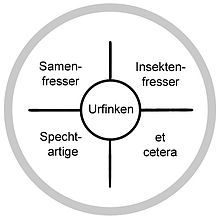 Schema der Radiation der Darwinfinken aus einem gemeinsamen Vorfahren
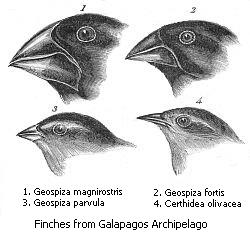
Schema der Radiation der Darwinfinken aus einem gemeinsamen Vorfahren Schnabelformen verschiedener Darwinfinken
Schnabelformen verschiedener DarwinfinkenUnter adaptiver Radiation (lateinisch: adaptare – anpassen; radiatus – strahlend) versteht man in der Evolutionsbiologie die Auffächerung (Radiation) einer wenig spezialisierten Art durch Herausbildung spezifischer Anpassungen (Adaptationen) an die vorhandenen Umweltverhältnisse in viele stärker spezialisierte Arten. Damit verbunden ist die Ausnutzung unterschiedlicher, vorher nicht besetzter ökologischer Nischen. Dieser Vorgang wird auch als „Allogenese“, „Allomorphose“, „Cladogenese“ oder „Idioadaptation“ bezeichnet.
Inhaltsverzeichnis
Artbildung in isolierten Lebensräumen
 Unterwasseraufnahme von Buntbarschen (Mbuna) im ostafrikanischen Malawisee. Die sehr artenreiche Buntbarschfauna dieses Großgewässers lässt sich auf eine kleine Gründerpopulation zurückführen, die sich durch Einnischung in einen Schwarm endemischer Arten aufgefächert hat.
Unterwasseraufnahme von Buntbarschen (Mbuna) im ostafrikanischen Malawisee. Die sehr artenreiche Buntbarschfauna dieses Großgewässers lässt sich auf eine kleine Gründerpopulation zurückführen, die sich durch Einnischung in einen Schwarm endemischer Arten aufgefächert hat.
Artbildung durch adaptive Radiation gilt als einer der grundlegenden Mechanismen der Evolution. Triebkräfte der adaptiven Radiation sind also wie bei allen Evolutionsvorgängen die genetische Variation und natürliche Auslese (Selektion; z. B. durch intraspezifische Konkurrenz[1]) innerhalb einer Population. Besonders eindrucksvolle Beispiele für adaptive Radiation ergeben sich bei der Besiedlung bisher unbesiedelter, isolierter Lebensräume, z. B. vulkanischer ozeanischer Inseln oder neu entstandener Süßwasserseen. Diese sind zunächst nur von wenigen Arten besiedelt, wodurch die einwandernden Arten viel weniger Konkurrenten haben als in „alten“ Lebensräumen mit vielen Arten und langer Evolutionsgeschichte. Hier kann man beobachten, dass manchmal in evolutionär kurzer Zeit (das sind einige tausend bis zehntausend Jahre) sehr viele neue Arten entstehen[2]. Dies gilt aber nur in den Fällen, in denen der Lebensraum neu entstanden ist. Zerfällt ein vorher zusammenhängender Lebensraum in getrennte Inseln, beobachtet man zwar auch mit der Zeit, wie sich die nun verinselten Populationen morphologisch auseinander entwickeln und neue Arten bilden, aber: mit beschleunigter Neubildung von Arten ist hier nicht zu rechnen. Entscheidend für die beschleunigte Artbildung scheint es zu sein, dass eine Art große Mengen von Ressourcen (insbesondere Nahrungsressourcen) ohne Konkurrenten zur Verfügung hat. Begünstigend für eine adaptive Radiation in einem Gebiet sind also unbesetzte ökologische Nischen.
Die Mechanismen der beschleunigten Artbildung bei der Besiedlung neuer Inseln (tatsächlicher Inseln oder Habitat-Inseln) stellen einen Testfall für die Evolutionstheorie dar und haben deshalb wissenschaftlich ganz besondere Beachtung gefunden.
Eine Art, die einen neu entstandenen Lebensraum besiedelt wird normalerweise relativ unspezialisiert sein. Dies liegt einerseits daran, dass Pionierarten generell weniger spezialisiert sind, andererseits benötigt ein Spezialist meist besondere Habitate oder Lebensgemeinschaften, die noch gar nicht vorhanden sein können. Die neu ankommende Art kann die vorkommenden Ressourcen also mit recht geringer Effizienz ausnutzen. Das ist zunächst nicht so wichtig, weil sie ja keine potenziell überlegenen Konkurrenten hat. Mit der Zeit wird sich die Art durch Konkurrenz der Artgenossen (intraspezifische Konkurrenz) so verändern, dass sie die vorhandenen Ressourcen besonders gut nutzen kann. Da sich ihre Merkmale verändern und kein Genaustausch mit der Spenderpopulation besteht, entwickelt sie sich nach und nach zu einer getrennten, neuen Art. Damit ist die Besonderheit einer Inselspezies relativ leicht erklärbar. Allerdings: Warum entwickelt sich nicht nur eine neue Art, sondern viele? Für die Beantwortung dieser Frage hat die Wissenschaft keine eindeutige Antwort. Verschiedene Forscher haben verschiedene Modelle entwickelt. Möglicherweise sind in unterschiedlichen Fällen unterschiedliche Modelle richtig.
Für die Beantwortung der Frage sind zwei Teilprobleme zu unterscheiden:
- Die Veränderung des Körperbaus und der Morphologie
- Die Aufspaltung einer Art in zwei getrennte Arten
Arten können sich morphologisch stark verändern, ohne aufzuspalten, aufgespaltene Artenpaare können sich wiederum in ihrer Lebensweise extrem ähnlich bleiben, manchmal gestaltlich sogar nahezu (Zwillingsarten) oder tatsächlich (kryptische Arten) ununterscheidbar sein. Es handelt sich also um zwei getrennte Probleme.
Das klassische Modell
Die Begründer der synthetischen Evolutionstheorie, besonders Ernst Mayr und Theodosius Dobzhansky entwickelten ein Modell, bei dem zur Trennung einer Art im Normalfall eine Auftrennung in geographisch getrennte Teil-Lebensräume notwendig ist (Allopatrische Artbildung). Gibt es nicht eine Insel, sondern eine Inselgruppe (z. B. die Galapagos-Inseln oder die Hawaii-Inseln) kann sich auf jeder Insel eine Art bilden, wodurch die Artenzahl des Archipels ansteigt. Ob die getrennten Inselpopulationen tatsächlich Arten darstellen, erweist sich allerdings erst dann, wenn sie erneut in Kontakt treten. Besiedeln später Individuen von einer Insel eine andere, die bereits von einer verwandten Art besetzt ist, gibt es verschiedene Möglichkeiten: a) Die Populationen vermischen sich wieder. b) Die Populationen bleiben getrennt. Nur im Fall b haben sich zwei neue Arten gebildet. Wenn nicht eine der beiden Arten die andere durch Konkurrenzausschluss verdrängt, leben nun zwei Arten auf einer Insel. Dieselbe Sequenz kann nun viele Male hintereinander ablaufen, wodurch die Artenzahl nach und nach ansteigt.
Das Modell erklärt einige Fälle von adaptiver Radiation sehr gut, z. B. die Darwinfinken auf den Galapagos-Inseln[3][4] oder die hohe Artenzahl von Eidechsen der Gattung Anolis auf den karibischen Inseln[5] (wobei die tatsächlichen Verhältnisse natürlich viel verwickelter sind als hier vereinfacht dargestellt).
Andere Fälle sind schwieriger damit zu erklären, z. B. die Radiation von hunderten von Buntbarsch-Arten in den großen ostafrikanischen Seen. Ernst Mayr sprach hier vom "Cichlidenproblem"[6]
Ökologische Artbildung
In den letzten Jahren haben zahlreiche Forscher neue Modelle entwickelt, die die Entstehung neuer Arten auch bei räumlichem Kontakt der Ausgangspopulationen plausibel erscheinen lassen[7][8]. Man spricht hier von sympatrischer oder parapatrischer Artbildung. Wesentliche Grundlagen ist eine Selektion, die nicht in eine, sondern in verschiedene Richtungen wirkt. Beispielsweise kann sich eine Art, für die mittelgroße Beutetiere optimal wären, in einem Lebensraum mit viel besonders kleinen und besonders großen Beutetieren wiederfinden. Eine ungeteilte Population ist hier in einem Minimum gefangen, da sich die Vorteile und Nachteile beider möglichen Spezialisierungen aufheben. Bei Auftrennung in zwei getrennte Populationen kann sich jede auf eine Beuteklasse spezialisieren, wodurch die Gesamtfitness zunimmt. Der Vorgang wird als "disruptive Selektion" bezeichnet. Damit sich bei disruptiver Selektion getrennte Arten bilden können, sind weitere Annahmen notwendig. Entweder die Arten entwickeln sich zwar räumlich benachbart, aber doch getrennt mit schmaler Hybridzone ("parapatrisch", dieses Modell wurde bereits von Charles Darwin als Gedankenmodell verfolgt) oder bei Paarungen bevorzugen die Sexualpartner gezielt Partner mit einer ähnlichen Merkmalsausprägung wie bei ihnen selbst (Assortative Paarung, engl.: "assortative mating"). Man spricht in diesen Fällen von ökologischer Artbildung, weil die Trennung der Populationen nicht auf geographischen, sondern auf ökologischen Faktoren beruht. Möglicherweise lässt sich so besser erklären, wie in einem einzelnen See beinahe zeitgleich extrem viele Fischarten entstehen können[9]. An der Artbildung sind real vermutlich zahlreiche weitere Faktoren beteiligt. Ein in der Botanik lange bekanntes Phänomen, welches in der Zoologie lange Zeit vernachlässigt worden ist, ist z. B. der Einfluss von Hybriden auf die Radiation[10].
Merkmalsverschiebung
Bei der ökologischen Artbildung ist es die Verschiedenheit der entstehenden Arten selbst, die die Artentrennung auslöst. Entstehen zwei Arten hingegen durch allopatrische Artbildung nach dem klassischen Modell, werden sie zwar normalerweise etwas verschieden sein, weil unterschiedliche Inseln unterschiedliche Selektionsbedingungen aufweisen, oder einfach aus Zufall ("Gründereffekt"). Die teils eindrucksvollen Anpassungen der Arten an unterschiedliche Lebensräume ("Einnischung") erklärt dies allein aber in der Regel nicht. Hier kommt es, wenn vorher getrennte Populationen später wieder in Kontakt kommen, oft zu einer Merkmalsverschiebung (engl.: "charakter displacement")[11][12]. Durch Merkmalsverschiebung werden vorher ähnliche Arten oder Populationen unähnlicher, weil auch auf sie eine disruptive Selektion einwirkt. Individuen mit besonders ähnlichen Merkmalen, die eine ähnliche Ernährungsweise besitzen, unterliegen einer stärkeren Konkurrenz. Durch Merkmalsverschiebung können sich verschiedene Arten einen Lebensraum untereinander aufteilen und so die zwischenartliche (interspezifische) Konkurrenz verringern. Manchmal entstehen in benachbarten Seen oder auf benachbarten Inseln unabhängig voneinander Gruppen von Arten mit jeweils exakt gleichen Spezialisierungen. Beobachtet wurde dies z. B. bei Buntbarscharten in ostafrikanischen Seen [13], bei Stichlingen in Seen in Nordamerika[14] oder, besonders eindrucksvoll, bei Spinnenarten auf den Hawaii-Inseln[15].
Der tatsächliche Nachweis einer Merkmalsverschiebung (im hier definierten Sinne) setzt den Nachweis voraus, dass die beteiligten Arten untereinander konkurrieren, und ist deshalb nicht leicht zu führen. Arten können sich in unterschiedlichen Lebensräumen natürlich auch einfach per Zufall in unterschiedliche Richtungen entwickeln (z. B. plausibel für einige Arten von Darwinfinken[16]).
Rolle der sexuellen Selektion
Mehrere gut belegte Fallbeispiele, z. B. Drosophila-Arten auf den Hawaii-Inseln oder Buntbarsche in den ostafrikanischen Seen, deuten auf eine große Bedeutung sexueller Selektion auf die Artentstehung in diesen Fällen hin[17][18]. Typisch sind Arten mit auffallend gefärbten oder gezeichneten Männchen und sehr viel ähnlicheren und unauffälligeren Weibchen (sexuell dimorphe Arten). Die Arten unterscheiden sich in der Färbung der Männchen und in der Präferenz der Weibchen für diese Färbungen, experimentell erzeugte Hybriden können unbeschränkt lebensfähig sein ("präzygotische" Isolation). Ob und in welchem Ausmaß sexuelle Selektion die sympatrische Artbildung signifikant erleichtert, ist in der Forschung noch umstritten. Ein möglicher Mechanismus der erleichterten Artbildung in diesen Fällen wurde durch die Theorie der "sexuell antagonistischen Koevolution" entwickelt, bei der die unterschiedlichen Interessen beider Geschlechter bei Kosten und Nutzen der Fortpflanzung ein koevolutionäres "Wettrüsten" bewirken können, durch das sich für die Paarung bedeutsame Merkmale in kurzer Zeit stark verändern können[19].
Andere Fälle von Radiationen
Das oben dargestellte Modell der adaptiven Radiation wird in der Evolutionsforschung bereits seit langer Zeit auch auf andere Fälle angewandt[20]. Im Unterschied zu der Besiedlung von Inseln handelt es sich dabei aber nicht um aktuell stattfindende Evolutionsvorgänge, sondern um die Interpretation alter Aufspaltungen, die nur aus der fossilen Überlieferung bekannt sind. Damit sind diese nur durch Plausibilitätssschlüsse, nicht aber durch direkte Experimente zu belegen. Die wichtigsten Fälle sind:
- Radiation in Folge eines Massenaussterbens. Sterben in Folge eines von außen einwirkenden, katastrophalen Ereignisses zahlreiche Arten, die vorher gut angepasst waren, aus, stehen für die Überlebenden auf einmal Lebensräume und Nischen offen, die vorher durch diese überlegenen Konkurrenten versperrt waren. Ein klassisches Beispiel wäre die Radiation der Säugetiere nach dem Aussterben der Dinosaurier.
- Radiation in Folge einer Schlüssel-Innnovation. Erwirbt eine Art infolge langsamer Evolution ein Merkmal, welches ihr die Nutzung völlig neuer Lebensräume und neuer Ressourcen ermöglicht, oder ermöglicht ihr die Innovation, eine Ressource auf eine völlig neue Art und Weise zu nutzen, kann dieses neue Schlüsselmerkmal Erfolg und Verbreitung der Art in kurzer Zeit enorm beschleunigen, wodurch eine Radiation in unbesetzte Nischen leicht folgen kann. Für die artenreichste Klasse der Insekten wurden als Schlüssel-Innovationen z.B. vorgeschlagen: Flugfähigkeit, Flügelgelenk zum Einklappen der Flügel über den Hinterleib, holometabole Verwandlung mit Imagines und Larven mit völlig unterschiedlichem Körperbau und unterschiedlicher Lebensweise. Jeder dieser "Erfindungen" könnte eine beschleunigte Artbildung in der neuen Gruppe gefolgt sein, worauf die extrem unterschiedlichen heutigen Artenzahlen der jeweiligen Gruppen hindeuten.
Fallbeispiele
Bei den realen untersuchten Fällen erwiesen sich die verschiedenen dargestellten Hypothesen als unterschiedlich erfolgreich. Vor allem die tatsächliche Bedeutung der ökologischen Artbildung ist in der Wissenschaft noch umstritten. Durch Fortschritte in der Sequenzierung der DNA (PCR) ist es heute viel einfacher geworden, Stammbäume von Artengruppen aufzustellen, die eine kürzlich zurückliegende adaptive Radiation ausgeprägt haben. Dadurch ist es leichter geworden, Hypothesen zu testen. Gerade die hohe Geschwindigkeit der Artbildung bei manchen Fällen von adaptiver Radiation stellt hier allerdings ein Problem dar, da es schwierig ist, entsprechend schnell reagierende DNA-Marker zu finden. Trotz dieser Probleme in Details erscheinen die Vorgänge bei einer adaptiven Radiation im Prinzip gut erklärbar.
Bekannte Beispiele umfassen
- Darwinfinken auf den Galapagos-Inseln
- Kleidervögel auf Hawaii
- Buntbarsche der Afrikanischen Großen Seen
- Tenreks und Lemuren auf Madagaskar
- Riesenkrabbenspinnen asiatischer Gebirge, z. B. Himalaya
- Anolis auf Jamaika
- Kegelschnecken
- Fruchtfliegen oder Taufliegen (Drosophilidae) auf Hawaii
- Beutelsäuger in Australien
Literatur
- Ulrich Kutschera: Evolutionsbiologie, 3. Auflage, Verlag Eugen Ulmer, Stuttgart (2008), ISBN 3-8252-8318-6
- Dolph Schluter: The Ecology of Adaptive Radiation. Oxford: Oxford University Press. (2000)
Einzelnachweise
- ↑ Brigitte Meinhard: Abiturprüfung Bayern Biologie GK. Stark Verlagsgesellschaft mbG & Co. KG, 2009, ISBN 9783894490966, S. 2004-4
- ↑ In Bakterienkulturen können morphologisch unterscheidbare Stämme binnen drei Tagen entstehen: Paul B. Rainey & Michael Travisano (1998): Adaptive radiation in a heterogeneous environment. Nature 394: 69-72
- ↑ Peter R. Grant & B. Rosemary Grant: Adaptive Radiation of Darwin's Finches. American Scientist 90(2): 130
- ↑ David Lack (1947): Darwin's Finches. Cambridge: Cambridge University Press
- ↑ Losos, J. B. (1990): A phylogenetic analysis of character displacement in Caribbean Anolis lizards. Evolution 44: 1189–1203.
- ↑ Mayr E (1984) Evolution of fish species flocks: a commentary. In: Echelle, A.A., Kornfield, I. (editors) Evolution of fish species flocks. University of Maine at Orono Press: 3–12.
- ↑ Sergey Gavrilets & Aaron Vose (2005): Dynamic patterns of adaptive radiation. Proceedings of the National Academy of Science (PNAS) 102(50): 18040-18045
- ↑ Ulf Dieckmann, Michael Doebeli, Johan A. J. Metz, Diethard Tautz (2004): Adaptive Speciation. Cambridge University Press. ISBN 0521828422
- ↑ Walter Salzburger & Axel Meyer (2004): The species flocks of East African cichlid fishes: recent advances in molecular phylogenetics and population genetics. Naturwissenschaften 91: 277-290.
- ↑ Ole Seehausen (2004): Hybridization and adaptive radiation. TREE Trends in Ecology and Evolution 19(4): 198-207
- ↑ William L.Brown Jr. & Edward O. Wilson (1956): Character displacement. Systematic Zoology 5:49–64.
- ↑ Dolph Schluter (2000): Ecological character displacement in adaptive radiation. American Naturalist 156 (Supplement): S4-S16
- ↑ Thomas D.Kocher (2004): Adaptive Evolution and explosive speciation: The Cichlid fish model. Nature Reviews.Genetics 5: 288-298
- ↑ Dolph Schluter (1996): Ecological Causes of Adaptive Radiation. American Naturalist 148 (Supplement): S40-S64
- ↑ Rosemary Gillespie (2004): Community assembly through adaptive radiation in Hawaiian spiders. Science 303: 356-359
- ↑ Peter R. Grant and B. Rosemary Grant: Unpredictable Evolution in a 30-Year Study of Darwin’s Finches. Science 296 (2002): 707-711
- ↑ Thomas D. Kocher: Adaptive evolution and explosive speciation: The Cichlid fish model. Nature Reviews. Genetics 5 (2004): 288-298.
- ↑ Ringo, J. M.: Why 300 species of Hawaiian Drosophila? The sexual selection hypothesis. Evolution 31 (1977): 694-696
- ↑ vgl.z. B.: Sergey Gavrilets & Takehiko I. Hayashi (2005): Speciation and sexual conflict. Evolutionary Ecology Volume 19, Number 2, 167-198.
- ↑ Walter Sudhaus (2004): Radiation within the framework of evolutionary ecology. Organisms Diversity & Evolution Volume 4, Issue 3: 127-134. doi:10.1016/j.ode.2004.04.001

Wikimedia Foundation.