- Phototrophie
-
 Die Rotalge Stylonema alsidii ist phototroph.
Die Rotalge Stylonema alsidii ist phototroph.
Phototrophie oder Fototrophie (von altgriechisch φῶς phos = das Licht + τροφή trophé = die Ernährung) bezeichnet die Nutzung von Licht als Energiequelle durch Lebewesen. Das Licht wird gebraucht, um den energiereichen chemischen Stoff Adenosintriphosphat (ATP) als Energieüberträger und kurzfristigen Energiespeicher zu synthetisieren. Mit dieser ATP-Synthese wandeln Lebewesen die Lichtenergie in chemische Energie.
Nur bestimmte Organismen können Lichtenergie für ihren Stoffwechsel direkt nutzen. Sie heißen phototrophe Organismen oder Phototrophe. Phototrophie ist sowohl unter Prokaryoten (Lebewesen mit Zellen ohne Zellkern) als auch unter Eukaryoten (Lebewesen mit Zellen mit Zellkern) verbreitet. Die phototrophen Prokaryoten werden stoffwechselphysiologisch in zwei Gruppen gegliedert. Die erste Gruppe betreibt Photosynthese. Sie nutzen Lichtenergie mit Hilfe von Chlorophyll-Pigmenten (Chlorophylle oder Bacteriochlorophylle). Die zweite Gruppe benutzt keine Chlorophyll-Pigmente. Sie betreibt demzufolge keine Photosynthese. Solche Prokaryoten nutzen stattdessen Lichtenergie mit Hilfe der anders gearteten Rhodopsin-Pigmente Bacteriorhodopsin, Proteorhodopsin oder Xanthorhodopsin. Die phototrophen Eukaryoten zeigen keine ähnliche stoffwechselphysiologische Vielfalt. Alle phototrophen Eukaryoten greifen für ihre Phototrophie auf Chlorophyll-Pigmente zurück. Sie betreiben demnach sämtlichst Photosynthese.
Inhaltsverzeichnis
Formen der Phototrophie
Um Phototrophie durchzuführen, benötigen Organismen ganz bestimmte Farbstoffe (Pigmente). Diese besonderen Pigmente sitzen in Biomembranen. Dort absorbieren sie Licht und machen die in ihm enthaltene Strahlungsenergie nutzbar. In phototrophen Organismen wurden bisher zwei verschiedene Klassen solcher Pigmente entdeckt. Chlorophylle (Chlorophylle, Bacteriochlorophylle) und Rhodopsine (Bacteriorhodopsin, Proteorhodopsin, Xanthorhodopsin).
Phototrophie mit Chlorophyllen: Lichtabhängige Reaktion der Photosynthese
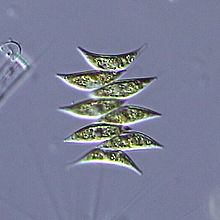 Die Grünalge Scenedesmus dimorphus wird phototroph durch Photosynthese.
Die Grünalge Scenedesmus dimorphus wird phototroph durch Photosynthese.Bei der Photosynthese wird Chlorophyll (oder Bacteriochlorophyll) durch Lichtenergie aus seinem chemischen Grundzustand in einen energiereichen („angeregten“) Zustand versetzt. Im angeregten Zustand gibt ein Chlorophyll-Molekül leicht ein energiereiches Elektron ab.
Das Elektron wird über bestimmte weitere Moleküle fort gereicht (Elektronentransportkette), die ebenfalls in der Biomembran sitzen. Im Zuge des Elektronentransports werden Wasserstoffkerne (Protonen, H+) von einer Seite der Biomembran auf die andere geschaufelt. Darum nimmt ihre Konzentration auf der einen Membranseite zu und gleichzeitig auf der anderen Seite ab. Auf diese Weise entsteht ein hohes H+-Konzentrationsgefälle zwischen beiden Membranseiten (Protonengradient).
Der Protonengradient wird genutzt, um ATP aufzubauen: In die Biomembran eingelassen ist die ATP-Synthase. Das ist ein Enzym, das die Synthese von ATP aus Adenosindiphosphat (ADP) und Phosphat (Pi) katalysiert. Innerhalb der ATP-Synthase verläuft ein Kanal, der beide Membranseiten verbindet. Durch den Kanal strömen Protonen entlang ihres Konzentrationsgefälles (→ Diffusion). Die kinetische Energie der hindurch strömenden Wasserstoffkerne wird von der ATP-Synthase zur ATP-Synthese verwendet, also in chemische Energie gewandelt (→ Chemiosmose).Lichtenergie wird genutzt, um einen Protonengradienten aufzubauen. Der Protonengradient wird genutzt, um ATP zu synthetisieren. Dieser phototrophe Vorgang gehört zur sogenannten lichtabhängigen Reaktion der Photosynthese.[1]
Die Anbindung von Pi an andere Stoffe wird Phosphorylierung genannt. Während der lichtabhängigen Reaktion wird ADP mit Hilfe von Licht zu ATP phosphoryliert. Demzufolge heißt der Vorgang Photophosphorylierung.
Es werden verschiedene Formen der Photosynthese unterschieden. Bei der oxygenen Photosynthese werden Wassermoleküle gespalten. Die Spaltung der Wassermoleküle benötigt ebenfalls Licht und Chlorophyll (→ Photolyse). Aus den gespaltenen Wassermolekülen wird der Elektronennachschub für die Elektronentransportkette gewonnen. Außerdem wird Sauerstoff frei. Bei der anoxygenen Photosynthese werden für den Elektronennachschub andere organische oder anorganische Stoffe herangezogen und eben kein Wasser. Hierzu wird kein Licht benötigt und es wird kein Sauerstoff frei.[2]
anorganische Elektronendon(at)oren der Photosynthese Elektronendon(at)or Photosynthese-Form Vorkommen Eisen-II-Ionen (Fe2+) anoxygene Photosynthese Purpurbakterien[3] Nitrit (NO2−) anoxygene Photosynthese Purpurbakterien[4] elementarer Schwefel (S0) anoxygene Photosynthese Purpurbakterien[5] Schwefelwasserstoff (H2S) anoxygene Photosynthese grüne Nichtschwefelbakterien,[6] grüne Schwefelbakterien,[7] Purpurbakterien[8] Thiosulfat (S2O32−) anoxygene Photosynthese Purpurbakterien[9] Wasser (H2O) oxygene Photosynthese Cyanobakterien,[10] Prochlorophyta,[11] phototrophe Eukaryoten[12] Wasserstoff (H2) anoxygene Photosynthese grüne Nichtschwefelbakterien[13] Phototrophie mit Rhodopsinen
 Haloarchaeen werden mit Bacteriorhodopsin phototroph.
Haloarchaeen werden mit Bacteriorhodopsin phototroph.Die phototrophe ATP-Synthese (Photophosphorylierung) mit Hilfe von Bacteriorhodopsin, Proteorhodopsin oder Xanthorhodopsin verläuft ebenfalls chemiosmotisch. Ein Rhodopsin besteht aus einem Eiweißstoff, der eine ganze Biomembran durchspannt (Transmembranprotein). In dem Eiweißstoff befindet sich ein Molekül namens Retinal.
Fällt Licht einer bestimmten Wellenlänge auf das Retinal, verändert das Molekül seine Gestalt. Dabei gibt es ein Proton an die Außenseite der Biomembran ab. Dem Retinal wird daraufhin von der Membraninnenseite ein neuer Wasserstoffkern zugeführt. Mit dem neuen Proton fällt das Molekül zurück in seine Ausgangsgestalt – bis es wieder von Licht getroffen wird, es seine Gestalt erneut ändert und ein weiteres Mal ein Proton an die Membranaußenseite abgibt. Auf diese Weise entsteht bei einer Rhodopsin-basierten Phototrophie ein hohes H+-Konzentrationsgefälle zwischen beiden Membranseiten (Protonengradient).
Der Protonengradient wird abgebaut, indem durch den Kanal einer ATP-Synthase Protonen entlang ihres Konzentrationsgefälles auf die Membraninnenseite zurück strömen. Die kinetische Energie der hindurch strömenden Wasserstoffkerne wird zur ATP-Synthese genutzt (→ Funktion des Bacteriorhodopsins).[14]Phototrophe Organismen
Phototrophe Prokaryoten
Verschiedene Prokaryoten haben verschiedene Formen der Phototrophie evolviert. Einerseits entwickelte sich die ATP-Synthese mit Hilfe der Pigmente Bacteriorhodopsin, Proteorhodopsin oder Xanthorhodopsin.[15][16] Andererseits evolvierte unabhängig davon die Phototrophie mit Hilfe von Chlorophyll-Pigmenten, die Photosynthese.[17] Für viele der phototrophen Prokaryoten ist Phototrophie nicht die einzige Möglichkeit des Energiestoffwechsels. Gerade bei Dunkelheit können sie eventuell auf verschiedene Wege der chemotrophischen Energiebereitstellung ausweichen.
Phototrophe Prokaryoten Prokaryoten Trophie Phototrophie-Form Cyanobakterien Photohydroautotrophie oxygene Photosynthese[18] grüne Nichtschwefelbakterien Photoorganoheterotrophie oder Photolithoautotrophie anoxygene Photosynthese des Typs II[19] grüne Schwefelbakterien Photolithoautotrophie oder Photoorganoheterotrophie anoxygene Photosynthese des Typs I[20], sogar an hydrothermalen Quellen der Tiefsee[21] Haloarchaeen Photoorganoheterotrophie Phototrophie mit Bacteriorhodopsin[22] Heliobakterien Photoorganoheterotrophie anoxygene Photosynthese des Typs I[23] Nichtschwefelpurpurbakterien Photolithoautotrophie oder Photoorganoheterotrophie anoxygene Photosynthese des Typs II[24][25][26] Thermoplasmaten Photoorganoheterotrophie Phototrophie mit Proteorhodopsin[27] α-Proteobakterien Photoorganoheterotrophie anoxygene Photosynthese des Typs II[28][29] oder Phototrophie mit Proteorhodopsin[30] Fulvimarina pelagi (ein α-Proteobakterium)
Photoorganoheterotrophie anoxygene Photosynthese des Typs II und Phototrophie mit Xanthorhodopsin[31][32] β-Proteobakterien Photoorganoheterotrophie anoxygene Photosynthese des Typs II[33] γ-Proteobakterien Photoorganoheterotrophie anoxygene Photosynthese des Typs II[34] oder Phototrophie mit Proteorhodopsin[35] Flavobakterien Photoorganoheterotrophie Phototrophie mit Proteorhodopsin[36][37] Prochlorophyten Photohydroautotrophie oxygene Photosynthese[38] Salinibacter ruber Photoorganoheterotrophie Phototrophie mit Xanthorhodopsin[39][40] Schwefelpurpurbakterien Photolithoautotrophie anoxygene Photosynthese des Typs II[41] Phototrophe Eukaryoten
Eukaryoten waren ursprünglich nicht phototroph. Einige Eukaryoten erlangten jedoch die Fähigkeit zur Phototrophie, indem sie Lebensgemeinschaften zu gegenseitigem Nutzen (Mutualismen) mit phototrophen Organismen eingingen. Solche Mutualismen entstanden mehrfach, zu verschiedenen Zeiten und unabhängig voneinander. Plastiden stellen die am weitesten fortgeschrittene Form des phototrophen Mutualismus dar (→ Endosymbiontentheorie).[42]
Die meisten der phototrophen Eukaryoten sind photohydroautotroph. Das heißt, dass sie Kohlenhydrate ausschließlich mit Licht, Wasser und Kohlendioxid synthetisieren. Diese phototrophen Eukaryoten betreiben oxygene Photosynthese mit Hilfe ihrer mutualistischen Partner. Die beiden einzigen bisher bekannten Ausnahmen von dieser Regel sind der Einzeller Oxyrrhis marina und die Hornisse Vespa orientalis. Jener Einzeller benutzt zur Phototrophie ein Rhodopsin-Pigment. Diese Form der Phototrophie ist zwar unter Prokaryoten weit verbreitet, unter Eukaryoten jedoch wahrscheinlich höchst selten. Zwar kommen Rhodopsine weitläufig unter Eukaryoten vor. Allerdings dienen sie hier gemeinhin nicht der Phototrophie, sondern der Rezeption.[43] Jene Hornisse andererseits scheint eine gänzlich neuartige Form der Phototrophie evolviert zu haben. Sie benutzt dazu den gelben Farbstoff Xanthopterin in ihrem Außenskelett.
Für einige der phototrophen Eukaryoten stellt die Phototrophie nicht die einzige Nährstoffquelle dar. Sie können sich darüber hinaus noch chemoorganoheterotroph ernähren. Das heißt, dass sie sich auch mit Nährstoffen versorgen können, indem sie andere Organismen ganz oder teilweise fressen. Derlei sich sowohl autotroph als auch heterotroph ernährende Lebensformen heißen Mixotrophe.
Phototrophe Eukaryoten 01: phototrophe Protisten, mehrzellige Algen und Landpflanzen[44][45] eukaryotische Großgruppe phototropher Mutualist Chlorarachniophyta komplexe Chloroplasten Chloroplastida: u.a. Chlorophyta (Grünalgen) und Charophyta mit Embryophyta (Landpflanzen) Chloroplasten Chrysophyta (Goldalgen): Bacillariophyceae (Kieselalgen), Chrysophyceae (Goldbraune Algen) und Xanthophyceae (Gelbgrüne Algen) komplexe Rhodoplasten Cryptophyta komplexe Rhodoplasten Dinophyta (Dinoflagellaten) komplexe Rhodoplasten Euglenophyta (Augentierchen) komplexe Chloroplasten Glaucophyta Cyanellen Haptophyta (Kalkalgen) komplexe Rhodoplasten Phaeophyta (Braunalgen) Phaeoplasten (komplexe Rhodoplasten) Rhodophyta (Rotalgen) Rhodoplasten Phototrophe Eukaryoten 02: Protisten mit besonderen Phototrophien[46] Protist eukaryotische Großgruppe besonderer phototropher Mutualist oder andere Form der Phototrophie
Auranticordis quadriverberis Cercozoa Cyanobakterien[47] Dinophysis spec. Dinophyta (Dinoflagellaten) komplexe Rhodoplasten aus Cryptophyta[48] oder Haptophyta (Kalkalgen)[49] Durinskia baltica Dinophyta (Dinoflagellaten) komplexe Rhodoplasten aus Bacillariophyceae (Kieselalgen)[50] Gymnodinium galatheanum Dinophyta (Dinoflagellaten) komplexe Rhodoplasten aus Haptophyta (Kalkalgen)[51] Gyrodinium aureolum Dinophyta (Dinoflagellaten) komplexe Rhodoplasten aus Haptophyta (Kalkalgen)[52] Großforaminiferen Foraminifera (Foraminiferen) Zoochlorellen oder Zooxanthellen[53] Hatena arenicola Cryptophyta Nephroselmis (Chlorophyta, Grünalgen)[54] Karenia brevis, Karenia digitata, Karenia mikimotoi
Dinophyta (Dinoflagellaten) komplexe Rhodoplasten aus Haptophyta (Kalkalgen)[55][56][57][58] Kryptoperidinium foliaceum Dinophyta (Dinoflagellaten) komplexe Rhodoplasten aus Bacillariophyceae (Kieselalgen)[59] Lepidodinium viride, Lepidodinium chlorophorum
Dinophyta (Dinoflagellaten) komplexe Chloroplasten aus Chlorophyta (Grünalgen)[60] Oxyrrhis marina Dinozoa Phototrophie mit Proteorhodopsin[61] Paramecium bursaria Ciliophora (Wimpertierchen) Zoochlorellen[62] Paulinella chromatophora Cercozoa Prochlorococcus oder Synechococcus (Cyanobacterien)[63][64] Phototrophe Eukaryoten 03: phototrophe Opisthokonten (Tiere und Pilze) Opisthokont phototropher Mutualist oder andere Form der Phototrophie
Ambystoma maculatum (Flecken-Querzahnmolch) Oophila amblystomatis (Chlorophyta, Grünalgen) in Eiern und Embryonen[65][66] Anthopleura elegantissima, A. xanthogrammica (Riesenanemonen)
Zoochlorellen und Zooxanthellen[67] Symsagittifera roscoffensis (grüner Acoelomorph) Tetraselmis convolutae (Chlorophyta, Grünalgen)[68] Hydra viridissima (Grüne Hydra) Zoochlorellen[69] Korallen Zooxanthellen[70][71] Lichenes (Flechten) Photobionten (Chlorophyta oder Cyanobakterien)[72] Sacoglossa (Schlundsackschnecken) phototrophe Plastiden aus gefressenen Chrysophyta (Goldalgen) oder Chlorophyta (Grünalgen) (→ Kleptoplastidie)[73] Tridacnidae (Riesenmuscheln) Zooxanthellen[74] Vespa orientalis (Orientalische Hornisse) Xanthopterin in der Cuticula[75] Siehe auch
Literatur
- Donat-Peter Häder (Hrsg.): Photosynthese. Stuttgart · New York, 1999 ISBN 3-13-115021-1
- Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006 ISBN 3-8273-7187-2
- Sonnewald U: Physiologie. In: Strasburger Lehrbuch der Botanik. Heidelberg, 2008 ISBN 978-3-8274-1455-7
Einzelnachweise
- ↑ Sonnewald U: Physiologie. In: Strasburger Lehrbuch der Botanik. Heidelberg, 2008: 274 ISBN 978-3-8274-1455-7
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 604-617 ISBN 3-8273-7187-2
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 613-614 ISBN 3-8273-7187-2
- ↑ Griffin BM, Schott J, Schink B: Nitrite, an electron donor for anoxygenic photosynthesis. In: Science 316 (2007): 1870 doi:10.1126/science.1139478
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 613-614 ISBN 3-8273-7187-2
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 621 ISBN 3-8273-7187-2
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 613-614 ISBN 3-8273-7187-2
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 456 ISBN 3-8273-7187-2
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 613-614 ISBN 3-8273-7187-2
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 444-447 ISBN 3-8273-7187-2
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 447 ISBN 3-8273-7187-2
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 448-449 ISBN 3-8273-7187-2
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 621 ISBN 3-8273-7187-2
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 478 ISBN 3-8273-7187-2
- ↑ Sharma AK, Spudich JL, Doolittle WF: Microbial rhodopsins: functional versatility and genetic mobility. In: Trends in Microbiology 14 (2006): 463-469 doi:10.1016/j.tim.2006.09.006
- ↑ Martinez A, Bradley AS, Waldbauer JR, Summons RE, DeLong EF: Proteorhodopsin photosystem gene expression enables photophosphorylation in a heterologous host. In: PNAS 104 (2007): 5590-5595 doi:10.1073/pnas.0611470104
- ↑ Bryant DA, Frigaard N-U: Prokaryotic photosynthesis and phototrophy illuminated. In: Trends in Microbiology 14 (2006): 488-496 doi:10.1016/j.tim.2006.09.001
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 444-447 ISBN 3-8273-7187-2
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 465 ISBN 3-8273-7187-2
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 456 ISBN 3-8273-7187-2
- ↑ Beatty JT, Overmann J, Lince MT, Manske AK, Lang AS, Blankenship RE, Van Dover CL, Martinson TA, Plumley FG: An obligately photosynthetic bacterial anaerobe from a deep-sea hydrothermal vent. In: PNAS 102 (2005): 9306-9310 doi:10.1073/pnas.0503674102 pdf
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 476,478 ISBN 3-8273-7187-2
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 431 ISBN 3-8273-7187-2
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 376 ISBN 3-8273-7187-2
- ↑ Duchow E, Douglas HC: Rhodomicrobium vannielii, a new photoheterotrophic bacterium. In: Journal of bacteriology 58 (1949): 409–416 pdf
- ↑ Straub KL, Rainey FA, Widdell F: Rhodovulum iodosum sp. nov. and Rhodovulum robiginosum sp. nov., two new marine phototrophic ferrous-iron-oxidizing purple bacteria. In: International Journal of Systematic Bacteriology 49 (1999): 729-735 pdf
- ↑ Frigaard NU, Martinez A, Mincer TJ, DeLong EF: Proteorhodopsin lateral gene transfer between marine planktonic Bacteria and Archaea. In: Nature 439 (2006): 847-850 doi:10.1038/nature04435
- ↑ Yurkov VV, Beatty JT: Aerobic Anoxygenic Phototrophic Bacteria. In: Microbiology and Molecular Biology Reviews 62 (1998): 695–724 pdf
- ↑ Jiao N, Zhang Y, Zeng Y, Hong N, Liu R, Chen F, Wang P: Distinct distribution pattern of abundance and diversity of aerobic anoxygenic phototrophic bacteria in the global ocean.. In: Environmental Microbiology 9 (2007): 3091-3099 doi:10.1111/j.1462-2920.2007.01419.x pdf
- ↑ de la Torre JR, Christianson LM, Béjà O, Suzuki MT, Karl DM, Heidelberg J, DeLong EF: Proteorhodopsin genes are distributed among divergent marine bacterial taxa. In: PNAS 100 (2003): 12830-12835 doi:10.1073/pnas.2133554100
- ↑ Cho JC, Giovannoni SJ: Fulvimarina pelagi gen. nov., sp. nov., a marine bacterium that forms a deep evolutionary lineage of descent in the order "Rhizobiales". In: International Journal of Systematic and Evolutionary Microbiology 53 (2003): 1853-1859 doi:10.1099/ijs.0.02644-0
- ↑ Kang I, Oh HM, Lim SI, Ferriera S, Giovannoni SJ, Cho JC: Genome Sequence of Fulvimarina pelagi HTCC2506T, a Mn(II)-Oxidizing Alphaproteobacterium Possessing an Aerobic Anoxygenic Photosynthetic Gene Cluster and Xanthorhodopsin. In: Journal of Bacteriology 192 (2010): 4798-4799 doi:10.1128/JB.00761-10
- ↑ Jiao N, Zhang Y, Zeng Y, Hong N, Liu R, Chen F, Wang P: Distinct distribution pattern of abundance and diversity of aerobic anoxygenic phototrophic bacteria in the global ocean.. In: Environmental Microbiology 9 (2007): 3091-3099 doi:10.1111/j.1462-2920.2007.01419.x pdf
- ↑ Jiao N, Zhang Y, Zeng Y, Hong N, Liu R, Chen F, Wang P: Distinct distribution pattern of abundance and diversity of aerobic anoxygenic phototrophic bacteria in the global ocean.. In: Environmental Microbiology 9 (2007): 3091-3099 doi:10.1111/j.1462-2920.2007.01419.x pdf
- ↑ Gómez-Consarnau l, Akram n, Lindell k, Pedersen A, Neutze R, Milton DL, González JM, Pinhassi J: Proteorhodopsin Phototrophy Promotes Survival of Marine Bacteria during Starvation. In: PLoS Biology 8 (2010): e1000358 doi:10.1371/journal.pbio.1000358
- ↑ Gómez-Consarnau L, González JM, Coll-Lladó M, Gourdon P, Pascher T, Neutze R, Pedrós-Alió C, Pinhassi J: Light stimulates growth of proteorhodopsin-containing marine Flavobacteria. In: Nature 445 (2007): 210-213 doi:10.1038/nature05381
- ↑ Riedel T, Tomasch J, Buchholz I, Jacobs J, Kollenberg M, Gerdts G, Wichels A, Brinkhoff T, Cypionka H, Wagner-Döbler I: Constitutive expression of the proteorhodopsin gene by a flavobacterium strain representative of the proteorhodopsin-producing microbial community in the North Sea. In: Applied and Environmental Microbiology 76 (2010): 3187-3197 doi:10.1128/AEM.02971-09
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 447 ISBN 3-8273-7187-2
- ↑ Antón J, Oren A, Benlloch S, Rodríguez-Valera F, Amann R, Rosselló-Mora R: Salinibacter ruber gen. nov., sp. nov., a novel, extremely halophilic member of the Bacteria from saltern crystallizer ponds. In: International Journal of Systematic and Evolutionary Microbiology 52 (2002): 485-91 pdf
- ↑ Lanyi JK, Balashov SP: Xanthorhodopsin: a bacteriorhodopsin-like proton pump with a carotenoid antenna. In: Biochimica et Biophysica Acta (BBA) - Bioenergetics 1777 (2008): 684-688 doi:10.1016/j.bbabio.2008.05.005
- ↑ Madigan MT, Martinko JM: Brock Mikrobiologie. München, 2006: 375 ISBN 3-8273-7187-2
- ↑ Keeling PJ: Diversity and evolutionary history of plastids and their hosts. In: American Journal of Botany (2004): 1481-1493 doi:10.3732/ajb.91.10.1481
- ↑ Deisseroth K: Lichtschalter im Gehirn. In: Spektrum der Wissenschaft 02/2011: 25
- ↑ Sonnewald U: Physiologie. In: Strasburger Lehrbuch der Botanik. Heidelberg, 2008: 224-225 ISBN 978-3-8274-1455-7
- ↑ Keeling PJ: Diversity and evolutionary history of plastids and their hosts. In: American Journal of Botany (2004): 1483-1487 doi:10.3732/ajb.91.10.1481
- ↑ Keeling PJ: Diversity and evolutionary history of plastids and their hosts. In: American Journal of Botany (2004): 1487 doi:10.3732/ajb.91.10.1481
- ↑ Chantangsi C, Esson HJ, Leander BS: Morphology and molecular phylogeny of a marine interstitial tetraflagellate with putative endosymbionts: Auranticordis quadriverberis n. gen. et sp. (Cercozoa). In: BMC Microbiology 8 (2008): 123 doi:10.1186/1471-2180-8-123
- ↑ Hackett JD, Maranda L, Yoon HS, Bhattacharya D: Phylogenetic evidence for the cryptophyte origin of the plastid of Dinophysis (Dinophysiales, Dinophyceae). In: Journal of Phycology 39 (2003): 440-448 doi:10.1046/j.1529-8817.2003.02100.x pdf
- ↑ Koike K, Sekiguchi H, Kobiyama A, Takishita K, Kawachi M, Koike K, Ogata T: A Novel Type of Kleptoplastidy in Dinophysis (Dinophyceae): Presence of Haptophyte-type Plastid in Dinophysis mitra. In: Protist 156 (2005): 225-237 doi:10.1016/j.protis.2005.04.002
- ↑ Imanian B, Pombert J-F, Keeling PJ: The Complete Plastid Genomes of the Two ‘Dinotoms’ Durinskia baltica and Kryptoperidinium foliaceum. In: PLoS ONE 5 (2010): e10711 doi:10.1371/journal.pone.0010711 pdf
- ↑ Tengs T, Dahlberg OJ, Shalchian-Tabrizi K, Klaveness D, Rudi K, Delwiche CF, Jakobsen KS: Phylogenetic Analyses Indicate that the 199Hexanoyloxy-fucoxanthin-Containing Dinoflagellates Have Tertiary Plastids of Haptophyte Origin. In: Molecular Biology and Evolution 17 (200): 718–729 pdf
- ↑ Tengs T, Dahlberg OJ, Shalchian-Tabrizi K, Klaveness D, Rudi K, Delwiche CF, Jakobsen KS: Phylogenetic Analyses Indicate that the 199Hexanoyloxy-fucoxanthin-Containing Dinoflagellates Have Tertiary Plastids of Haptophyte Origin. In: Molecular Biology and Evolution 17 (200): 718–729 pdf
- ↑ Blank RJ, Trench RK: Nomenclature of Endosymbiotic Dinoflagellates. In: Taxon 35 (1986): 386-394
- ↑ Okamoto N, Inouye I: Hatena arenicola gen. et sp. nov., a katablepharid undergoing probable plastid acquisition. In: Protist 157 (2006): 401-419 doi:10.1016/j.protis.2006.05.011
- ↑ Tengs T, Dahlberg OJ, Shalchian-Tabrizi K, Klaveness D, Rudi K, Delwiche CF, Jakobsen KS: Phylogenetic Analyses Indicate that the 199Hexanoyloxy-fucoxanthin-Containing Dinoflagellates Have Tertiary Plastids of Haptophyte Origin. In: Molecular Biology and Evolution 17 (200): 718–729 pdf
- ↑ Yang ZB, Takayama H, Matsuoka K, Hodgkiss IJ: Karenia digitata sp. nov. (Gymnodiniales, Dinophyceae), a new harmful algal bloom species from the coastal waters of west Japan and Hong Kong. In: ''Phycologia 39 (2000): 463-470 doi:10.2216/i0031-8884-39-6-463.1
- ↑ Hansen G, Daugbjerg N, Henriksen P:Comparative study of Gymnodinium mikimotoi and Gymnodinium aureolum, comb. nov. (=Gyrodinium aureolum) based on morphology, pigment composition, and molecular data. In: Journal of Phycology 36 (2000): 394–410 doi:10.1046/j.1529-8817.2000.99172.x pdf
- ↑ Daugbjerg N, Hansen G, Larsen L, Moestrup Ø: Phylogeny of some of the major genera of dinoflagellates based on ultrastructure and partial LSU rDNA sequence data, including the erection of three new genera of unarmoured dinoflagellates. In: Phycologia 39 (2000): 302-317 doi:10.2216/i0031-8884-39-4-302.1 pdf
- ↑ Imanian B, Pombert J-F, Keeling PJ: The Complete Plastid Genomes of the Two ‘Dinotoms’ Durinskia baltica and Kryptoperidinium foliaceum. In: PLoS ONE 5 (2010): e10711 doi:10.1371/journal.pone.0010711 pdf
- ↑ Hansen G, Botes L, Salas MD: Ultrastructure and large subunit rDNA sequences of Lepidodinium viride reveal a close relationship to Lepidodinium chlorophorum comb. nov. (= Gymnodinium chlorophorum). In: Phycological Research 55 (2007): 25-41 doi:10.1111/j.1440-1835.2006.00442.x pdf
- ↑ Slamovits CH, Okamoto N, Burri L, James ER, Keeling PJ: A bacterial proteorhodopsin proton pump in marine eukaryotes. In: Nature Communications 2 (2010): 183 doi:10.1038/ncomms1188
- ↑ Karakashian MW: Symbiosis in Paramecium Bursaria. In: Symposia of the Society of Experimental Biology 29 (1975): 145-73 abstract
- ↑ Yoon HS, Nakayama T, Reyes-Prieto A, Andersen RA, Boo SM, Ishida K, Bhattacharya D: A single origin of the photosynthetic organelle in different Paulinella lineages. In: BMC Evolutionary Biology 9 (2009): 98 doi:10.1186/1471-2148-9-98
- ↑ Nakayama T, Ishida K-i: Another acquisition of a primary photosynthetic organelle is underway in Paulinella chromatophora. In: Current Biology 19 (2009): R284-R285 doi:10.1016/j.cub.2009.02.043 pdf
- ↑ Gilbert PW: The alga-egg relationship in Ambystoma maculatum, a case of symbiosis. In: Ecology 25 (1944): 366-369 doi:10.2307/1931284
- ↑ Kerney R, Kim E, Hangarter RP, Heiss AA, Bishop CD, Hall BK: Intracellular invasion of green algae in a salamander host. In: PNAS 108 (2011): 6497-6502 doi:10.1073/pnas.1018259108
- ↑ Secord D, Augustine L: Biogeography and microhabitat variation in temperate algal-invertebrate symbioses: zooxanthellae and zoochlorellae in two Pacific intertidal sea anemones, Anthopleura elegantissima and A. xanthogrammica. In: Invertebrate Biology 119 (2000): 139-146 doi:10.1111/j.1744-7410.2000.tb00002.x
- ↑ Douglas AE: Growth and Reproduction of Convoluta Roscoffensis Containing Different Naturally Occurring Algal Symbionts. In: Journal of the Marine Biological Association of the United Kingdom 65 (1985): 871-879 doi:10.1017/S0025315400019378
- ↑ Dunn K: Growth of endosymbiotic algae in the green hydra, Hydra viridissima. In: Journal of Cell Science 88 (1987): 571-578 pdf
- ↑ Rowan R Knowlton N: Intraspecific diversity and ecological zonation in coral-algal symbiosis. In: PNAS 92 (1995): 2850-2853 pdf
- ↑ Little AF, van Oppen MJ, Willis BL: Flexibility in algal endosymbioses shapes growth in reef corals. In: Science 304 (2004): 1492-1494 doi:10.1126/science.1095733
- ↑ Rambold G, Friedl T, Beck A: Photobionts in lichens: Possible indicators of phylogenetic relationships? In: The Bryologist 101 (1998): 392–397
- ↑ Hinde R, Smith DC: “Chloroplast symbiosis” and the extent to which it occurs in Sacoglossa (Gastropoda: Mollusca). In: Biological Journal of the Linnean Society 6 (1974): 349-356 doi:10.1111/j.1095-8312.1974.tb00729.x
- ↑ Trench RK, Wethey DS, Porter JW: Observations on the symbiosis with zooxanthellae among the Tridacnidae (Mollusca, Bivalvia). In: Biological Bulletin 161 (1981): 180-198 pdf
- ↑ Plotkin M, Hod I, Zaban A, Boden SA, Bagnall DM, Galushko D, Bergman DJ: Solar energy harvesting in the epicuticle of the oriental hornet (Vespa orientalis). In: Naturwissenschaften 97 (2010): 1067-1076 doi:10.1007/s00114-010-0728-1
Kategorien:- Zellbiologie
- Ökologische Eigenschaft
- Ernährungstyp


Wikimedia Foundation.