- Flügel (Insekt)
-
Die Beschäftigung mit dem Insektenflügel gehört zu den zentralen Themen der Entomologie, der Lehre von den Insekten. Das Verständnis seiner Entstehung und seiner Formenvielfalt stellt eine große Herausforderung für viele biologische Teildisziplinen dar.
Für die Benennung der Adern, Zellen und Felder des Flügels existieren verschiedene Systeme. Die Evolution der Flügel ist weitgehend ungeklärt und auch bezüglich der Entwicklung während der Ontogenese (Individualentwicklung) sind noch viele Fragen unbeantwortet.
Bereits im frühen Oberkarbon und damit mehr als 100 Millionen Jahre vor den Flugsauriern, den ersten flugfähigen Landwirbeltieren, die mit zu Flügeln umgebildeten Gliedmaßen zu fliegen vermochten, entwickelten die Insekten Flügel aus Ausstülpungen der Haut. Die Entwicklung der Flugfähigkeit ermöglichte die Eroberung neuer Lebensräume und die Besetzung von zahlreichen neuen ökologischen Nischen. In Folge davon kam es zu einer umfangreichen adaptiven Radiation, wodurch die Fluginsekten heute die artenreichste Tiergruppe überhaupt sind. Der Erfolg der Insekten ist auf die Entwicklung der Flügel zurückzuführen.
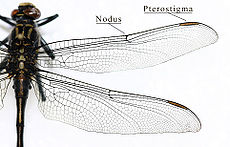

Abb. 0.1 Ursprüngliche Aderung und Flügelhaltung bei einer Großlibelle Abb. 0.2 Schwebfliegen bei der Paarung 

Abb. 0.3 Ausschnitt eines Schmetterlingsflügels mit Vergrößerung Abb. 0.4 Gehärtete Vorderflügel und faltbare Hinterflügel beim Maikäfer Inhaltsverzeichnis
Evolution
Theorien zur Entstehung
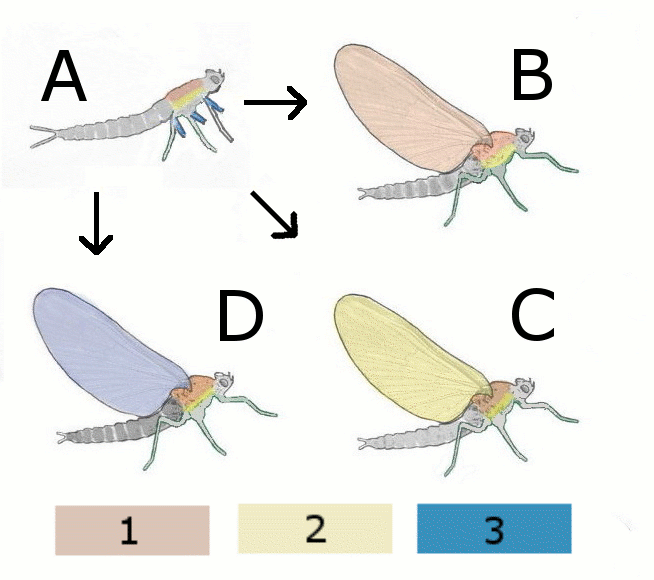
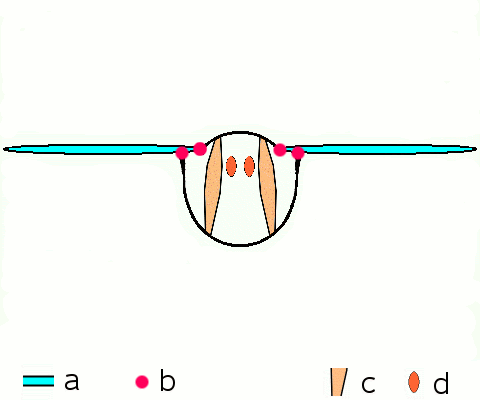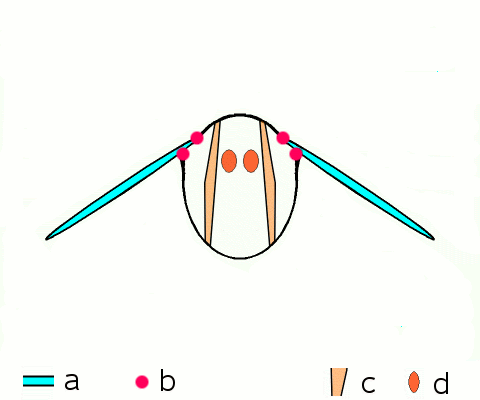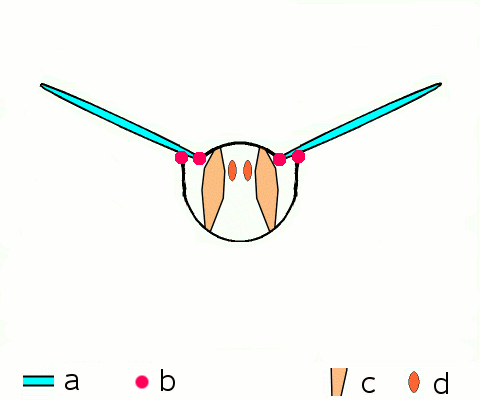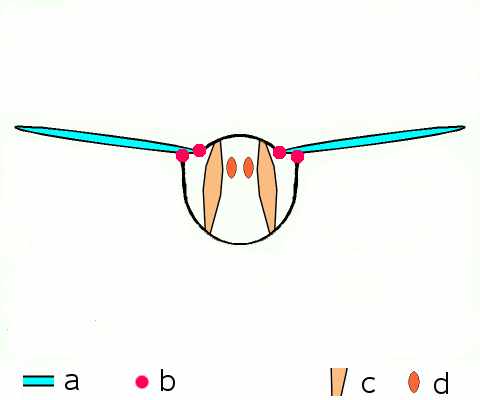
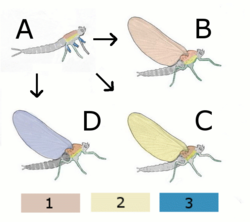 Abb. 1.1 Theorien zur Flügelentstehung
Abb. 1.1 Theorien zur Flügelentstehung
A Hypothetischer flügelloser Vorfahre
B Paranotaltheorie
Hypothetisches Insekt mit Flügel aus Rücken (Notum)
C Hypothetisches Insekt mit Flügel aus Seite (Pleurum)
D Epicoxal-Theorie
Hypothetisches Insekt mit Flügel aus Anhang der Beine
1 Notum (Rücken)
2 Pleurum (Seite)
3 Exiten (äußere Anhänge der Beine)Aus dem Devon (vor 400 Millionen Jahren) sind lediglich Fossilien flügelloser Urinsekten, aus dem Karbon (vor 320 Millionen Jahren) bereits Insekten aus mehr als 10 verschiedenen Gattungen mit voll funktionsfähigen Flügeln bekannt. Aus der Zeit dazwischen fehlen Funde, an denen Übergangsformen zum funktionierenden Flügel erkennbar wären.
So sind die Annahmen über den Verlauf der Evolution der Flügel weitgehend hypothetisch. Die Theorien, aus welchem Teil der Segmente der Brust sich die Flügel entwickelt haben, lassen sich in drei Gruppen gliedern (Abb. 1.1):
- Herkunft aus seitlichen Auswüchsen des Insekts,
- Ableitung aus Auswüchsen am Rücken des Insekts, und
- Entwicklung aus Anhängen der Beine des Insekts (Exiten- und Enditentheorien).
Heute werden hauptsächlich zwei Theorien diskutiert.
- Nach der Epicoxal-Theorie (Kukalová-Peck) sind die Flügel aus beweglichen seitlichen Anhängen an der Basis der Beine entstanden. Die Vorfahren der Insekten hatten demnach gegliederte Beine mit äußeren und inneren Anhängen. Aus diesen Beinen entwickelten sich einerseits die Spaltbeine der Krebse, andrerseits aus dem beweglichen äußeren Anhang des obersten Beingliedes, der Epicoxa, die Flügel der Insekten. Die Epicoxal-Theorie stützt sich dabei hauptsächlich auf die Tatsache, dass bei allen Insekten die Flügel durch eine Abzweigung der Beintrachee mit Sauerstoff versorgt werden, und auf paläontologische Hinweise.
- Nach der Paranotaltheorie (erstmals Müller, F., 1873[1]) haben sich die Flügel aus starren Auswüchsen der Rückenplatten (Notum) der Brust gebildet. Dafür spricht, dass während der Ontogenese von Insekten mit unvollständiger Entwicklung die Flügelanlagen bis zur letzten Häutung starr mit diesem Körperteil verbunden sind.
Unabhängig davon, ob die Flügel aus Anhängen der Beine oder Ausstülpungen der Brust entstanden sind, bleibt die Frage ungeklärt, weshalb sich diese Flügelvorläufer zum funktionsfähigen Flügel weiterentwickelt haben. Es müssen Gründe angegeben werden, weshalb es einen Selektionsvorteil bedeutete, dass anfängliche Flügelstummel sich so vergrößerten, dass der Funktionswechsel vom flugunfähigen Fortsatz zum flugfähigen Organ möglich war. Hierfür existieren mehrere plausible Hypothesen, die hier zusammenfassend aufgeführt werden.
- Die Körperauswüchse wurden zur Werbung eingesetzt. Größere Flügel könnten zu einer höheren Fortpflanzungswahrscheinlichkeit führen.
- Die Vorläufer der Flügel hatten direkt Kiemenfunktion oder sorgten über eine Ventilation des Wassers indirekt zu einer besseren Sauerstoffversorgung. Ihre ursprüngliche Beweglichkeit führte über Ruderfunktion und flugähnliches Rudern auf der Wasseroberfläche (surface-skimming) zur Flügelvergrößerung, gekoppelt mit der Entwicklung von Muskulatur und Gelenkigkeit.
- Die Vorformen der Flügel hatten eine Schutzfunktion für die Kiemen. Ihre Größe verbesserte den Schutz der Kiemen und wurde deswegen positiv selektioniert.
- Die Körperauswüchse boten aerodynamische Vorteile. Sie ermöglichten etwa beim Jagen der Beute oder bei der Flucht ein höheres oder weiteres Springen oder brachten eine kleinere Verletzungsgefahr beim Fallen aus großer Höhe mit sich. Schon eine höhere Wahrscheinlichkeit, beim Fallen gleich auf den Beinen zu landen, würde einen Selektionsvorteil bedeuten. So konnte über das Gleiten das Fliegen entwickelt werden.
- Die Vorformen der Flügel dienten der Wärmeregulation. Insekten als wechselwarme Tiere sind bei kühler Umgebung lethargisch. Es ist für sie von Vorteil, wenn sie bei Erwärmung früher die für die Beweglichkeit notwendige Temperatur erreichen als die anderen wechselwarmen Tiere. Die der Sonne ausgesetzten Vorformen der Flügel sollen mit einem dichten Netz von Adern durchzogen gewesen sein. Die Wärme, die durch Sonnenbestrahlung aufgenommen wurde, könnte so effizient in den Körper transportiert worden sein. In den Adern der Flügel gespeicherte Wärme würde auch eine Verlängerung des Aktivitätszeitraums bei Abkühlung ermöglichen. Die einfachste Art, die Körperoberfläche zu vergrößern und gleichzeitig das zu erwärmende Volumen klein zu halten, sind Auswüchse der Körperoberfläche in Form dünner Platten, die im Idealfall bei Erwärmung entfaltet, bei Abkühlung dicht am Körper gehalten werden und bei Entfaltung jeweils so gedreht und geneigt werden können, dass sie in günstigem Winkel zur Sonneneinstrahlung stehen.
Zur Abwägung dieser Hypothesen müsste zumindest etwas über die fragliche Insektengruppe, bei der die Entwicklung der Flügel einsetzte, bekannt sein. Je nachdem, ob es sich um ein Wasser- oder Landtiere gehandelt hat, wird die eine oder andere Hypothese plausibler, je nach Größe haben die aerodynamischen Überlegungen mehr Gewicht, je nach Lebensweise und Fortpflanzungsbiologie müssen die Hypothesen verschieden bewertet werden.
Abb. 1.2 Evolution der Möglichkeiten, die Flügel in Ruhestellung an den Körper anzulegen Flügel nicht nach hinten klappbar
(rezente Archaeoptera)seitlich ausgebreitet (Großlibellen) über dem Rücken aneinanderliegend
(Kleinlibellen, Eintagsfliegen)Klappbar
(Neoptera)Flügel nicht faltbar (z. B. Steinfliegen) Faltbar Längsfaltung (z. B..Vorderflügel der Faltenwespen, Abb. 1.3) Querfaltung (z. B. Hinterflügel der Käfer, Abb. 1.4) Fächerfaltung (z. B. Hinterflügel der Ohrwürmer, Abb. 1.5) Fossile Funde der Gattungen Stenodictya aus dem Karbon und Lemmatophora aus dem Perm (vor 270 Millionen Jahren) zeigen neben einfachen Flügeln am mittleren und hinteren Brustabschnitt flügelstummelähnliche seitlich abstehende Auswüchse am ersten Brustsegment. Diese früher als reduziertes drittes Flügelpaar angesehenen Stummel finden heute mit mehreren der oben genannten Hypothesen eine weit schlüssigere Erklärung (Bild bei Weblinks). In allen Fällen ist eine schnelle Zunahme der Flügelgröße und -beweglichkeit zu erwarten. Genügend große Auswüchse würden die Möglichkeit des Gleitfluges und der Verdriftung durch Wind ermöglichen und somit den Funktionswechsel zu Fortbewegungsorganen einleiten.[2][3][4][5][6][7][8]
Um die Herkunft der Insektenflügel zu rekonstruieren, haben kürzlich Wissenschaftler der japanischen Forschergruppe um Shigeo Hayashi die Expression von drei für die Flügelausformung entscheidenden Regulatorgenen bei Vertretern zweier Insektengruppen studiert, nämlich bei geflügelten, aber sehr ursprünglichen Eintagsfliegen (Ephemeroptera) und bei Silberfischchen, also ungeflügelten Urinsekten der Ordnung Zygentoma. [9]
Verschiedene Körperanhänge dieser ursprünglichen Gruppen wurden bereits vorher als Vorläufer der Flügel in Betracht gezogen. Die Ergebnisse lassen sich so interpretieren, dass bei den ungeflügelten Urinsekten zwei getrennte, entwicklungssteuernde Module präsent sind. Das eine wirkt auf die Körperseiten (lateral) ein und steuert die Ausbildung des oberen, stäbchenförmigen Asts der verzweigten Gliedmaßen. Das andere wirkt auf die Nahtstelle zwischen Körperseite und Rücken und steuert dort die Ausbildung von flachen, schildförmigen Auswüchsen.
Die Autoren schlagen ein Modell vor, nach dem die oberen (dorsalen) Äste der Gliedmaßen in den Bereich der Nahtstelle zwischen Körperseite und Rücken einbezogen wurden, und die Form der dort vorkommenden, flachen Auswüchse annahmen. Das geschah durch die einfache Integration der beiden vorher getrennten Steuer-Module, so dass anschließend sowohl die flache, zweischichtige Bauform der Flügel verfügbar wurde, als auch die (zu den Gliedmaßen gehörige) Muskulatur, die Bewegungen ausführen konnte. Dadurch wurde eine rasche Evolution der Insektenflügel möglich. Das vorliegende Modell zeigt, dass sich selbst große Bauplanänderungen, die eine enorme Auswirkung auf die Ökologie und weitere Stammesgeschichte einer Tiergruppe haben, auf der Ebene der Entwicklungssteuerung durch relativ wenige Veränderungsschritte erklären lassen.[10]
Die Entdeckung der fossilen Larven der Coxoplectoptera lieferte 2011 neue Hinweise zur Klärung des evolutionären Ursprungs der Insektenflügel, die zu einer ähnlichen Hypothese, wie derjenigen der Hayashi-Arbeitsgruppe führte. Bislang galten die zwei Theorien, Paranotal-Theorie und Exit-Theorie, als unvereinbare Alternativen, für die jeweils unterschiedliche Evidenzen aus Fossilbericht, vergleichender Morphologie, Entwicklungsbiologie und Genetik sprachen. Der Nachweis, dass Beingene bei der Ontogenese der larvalen Flügelanlagen exprimiert werden, galt als überzeugender Beleg für die Exit-Theorie, die den Insektenflügel von umgewandelten, beweglichen Ästen (Exiten) der Spaltbeine herleitet. Die Larven der Coxoplectoptera zeigen jedoch, dass die Hinterleibskiemen der Eintagsfliegen und ihrer Ahnen, die als den Flügeln seriell entsprechende Organe gelten, innerhalb der Rückenschilde entspringen. Bei modernen Eintagsfliegen ist dies nicht erkennbar, da im Hinterleib der Larven die Rücken- und Bauchschilde stets zu Ringen verwachsen sind, und auch embryonal keine Hinweise zu finden sind. Wenn Larvenkiemen und Flügel sich entsprechende (seriell homologe) Strukturen sind und somit gleichen evolutionären Ursprung haben, so bedeutet der Befund der Coxoplectoptera-Larven, dass auch die Flügel tergalen Ursprungs sind, wie es die klassische Paranotal-Theorie besagte. Staniczek, Bechly & Godunko (2011)[11] schlugen daher eine neue Hypothese vor, die die neuen paläontologischen Befunde mit den Ergebnissen der Entwicklungsgenetik in Einklang bringen könnte. Demnach seien Flügel zunächst als starre Auswüchse der Rückenschilde (Paranota) entstanden, und erst später in der Evolution wären diese Auswüchse durch die sekundäre "Rekrutierung" von Beingenen beweglich geworden.
Paläontologische Befunde
Obwohl eindeutige Zwischenstufen von geflügelten und ungeflügelten Insekten bislang nicht gefunden wurden, was von Gegnern der Evolutionstheorie immer wieder betont wird, lässt sich innerhalb der geflügelten Insekten eine Entwicklung durch Fossilien belegen. Das älteste bisher gefundene geflügelte Insekt ist etwa 324 Millionen Jahre alt benannt nach seinem Fundort im sächsischen/sachsen-anhaltischen Bergbaugebiet Bitterfeld/Delitzsch Delitzschala bitterfeldensis (Bild bei Weblinks). Außerdem befindet sich in Deutschland bei Hagen-Vorhalle (Westfalen) ein Fundort für fossile Insekten. Fundstätten sind über die ganze Welt verteilt, ein Hinweis auf eine Serie von acht Weltkarten aus acht Erdzeitaltern mit der Lage von Funden aus diesen Zeiten befindet sich im Absatz Weblinks.
Abb. 1.3 Längsfaltung am Beispiel der Faltenwespen (Vespidae) 
Im oberen Bild ist die Hauptfaltungslinie des Vorderflügels etwa auf halber Höhe als helle waagrechte Linie erkennbar. Der Flügelteil, der sich hinter dieser Linie befindet, wird nach hinten unten umgeschlagen. Der schmale Streifen am Vorderrand des Flügels vor der ersten starken Ader wird nach vorne unten umgeschlagen. 
So bildet in Ruhestellung die Ader den widerstandsfähigen Außenrand des Flügels, der die Seiten des Hinterleibs wie eine Art Stoßdämpfer schützt. Der Hinterflügel ist durch den Vorderflügel weitgehend umhüllt. Angesichts der grundsätzlichen Schwierigkeiten der Fossilisation so zerbrechlicher Lebewesen in alten Gesteinsschichten ist jedoch zu erwarten, dass viele Funde einen Interpretationsspielraum zulassen. Eine weitere grundsätzliche Schwierigkeit besteht darin, dass eine fossil belegte Neuerung in der davor liegenden Zeit entwickelt worden sein muss. So werden die Funde ergänzt durch zeitliche Abschätzungen mit der molekularen Uhr. Nach diesen liegt das Auftreten der ersten Fluginsekten im Devon vor ungefähr 390 Millionen Jahren. Die Aufspaltung in Palaeoptera und Neoptera wird im mittleren Devon vermutet.
Die Neoptera (Neuflügler) umfassen alle heutigen Insektenordnungen mit Flügeln außer den Eintagsfliegen und Libellen sowie die Insektenordnungen, bei denen die Flügel wieder zurückgebildet wurden. Sie besitzen einen einheitlichen Mechanismus, die Flügel nach hinten umzulegen, weshalb ein gemeinsamer Vorfahre angenommen wird (monophyletische Entstehung). Eine Radiation der Neoptera erfolgte vermutlich im oberen Karbon, da im Perm bereits die Mehrheit der heutigen Ordnungen erkennbar ist. Lediglich hochspezialisierte Insektenordnungen traten erst später auf. So sind zum Beispiel die Flöhe mit rückgebildeten Flügeln erst in der unteren Kreidezeit nachzuweisen.
Bei den Palaeoptera (Altflüglern, nach anderen Autoren Palaeopteroidea) mit etwa sieben Ordnungen, von denen fünf ausgestorben sind, sind die systematischen Zusammenhänge noch unklar. Bezüglich der Flügel sind drei Gruppen von Interesse.
Abb. 1.4 Querfaltung beim Rosenkäfer 
Hinterflügel ausgebreitet. Durch Faltungslinien ist er in fünf Felder unterteilt, die jeweils nach hinten durch einen Bogen abgeschlossen sind. 
Der gleiche Flügel halb gefaltet. Die beiden Gelenke für die Querfaltung bilden einen stumpfen Winkel. Rechts davon ist der Flügel bereits in drei Lagen gefaltet. Bei größerer Auflösung ist der dritte Bogen des Flügelrandes unter dem ersten und zweiten sichtbar. Links davon erscheint der fünfte Bogen unter dem vierten. 
Der gleiche Flügel vollständig gefaltet. Die fünf Felder liegen übereinander. (Der Vorderflügel wurde entfernt) - Bei einer Ordnung der Palaeoptera, den ausgestorbenen Diaphanopter(oide)a, findet sich ein altertümlicher Typ der Klappung der Flügel nach hinten. Dies zeigt, dass die Möglichkeit der Flügelklappung bei Ruhestellung unabhängig voneinander (polyphyletisch) im Karbon zweimal entwickelt wurde. Der Gedanke liegt nahe, dass diese Klappung einen Selektionsvorteil bedeutete oder anders ausgedrückt, dass nicht klappbare Flügel für gewisse Lebensräume und -formen nachteilig waren.
- Außer den Eintagsfliegen sind die Libellen die einzige Ordnung heutiger geflügelter Insekten, die nicht den Neoptera, sondern den Palaeoptera zugerechnet werden und bei denen in Ruhestellung keine Klappung der Flügel nach hinten möglich ist. Zu ihren Vorfahren gehört die Urlibelle Meganeuropsis permiana mit einer Flügelspannweite von 71 cm aus dem Perm. Außerdem existieren Fossilien von paläozoischen Urlibellenlarven, deren Flügelanlagen so gut ausgebildet sind, dass man die Flugfähigkeit der Larven vermuten kann. Auch andere paläozoische Fluginsekten besaßen schon in den frühesten bekannten Larvenstadien voll gelenkige Flügelscheiden, die oft seitlich abgespreizt ausgewachsen sind, im Gegensatz zu den starr und angelegt auswachsenden Flügelscheiden moderner Fluginsektenlarven. Die Flügelscheiden dieser paläozoischen Fluginsektenlarven waren bei frühen Larvenstadien deutlich gebogen, und bei späteren Larvenstadien wurden sie, neben der Größenzunahme, auch immer gerader und imaginalähnlicher.
- Bei den heutigen Vertretern der Eintagsfliegen sind die Hinterflügel deutlich kleiner als die Vorderflügel. Bei fossilen Funden findet man Formen, bei denen die Flügel noch gleiche Größe besitzen. Außerdem findet man bei den heutigen Eintagsfliegen, die zu den ursprünglichsten Fluginsekten zählen, als einzigen Fluginsekten das interessante Phänomen, dass neben der eigentlichen Imago auch das letzte "Larvenstadium" (Subimago), das lediglich eine Übergangsform darstellt, noch flugfähig ist und somit noch eine letzte Häutung im flugfähigen Zustand erfolgt. Es könnte daher sein, dass eine Begrenzung der Flugfähigkeit auf das vermehrungsfähige Adultstadium, wie sie heute bei allen übrigen Fluginsekten zu finden ist, einen Selektionsvorteil hatte.
Im Gegensatz zum Umklappmechanismus der Flügel bei den Diaphanoptera ermöglichte der Klappungstyp der Neoptera eine solche Beweglichkeit gewisser Flügeladern gegeneinander, dass in der Ruhestellung eine Längsfaltung der Flügel möglich wird. Sie bezieht sich in der Regel nur auf die Hinterflügel, lediglich bei den Hautflüglern auch auf die Vorderflügel (Abb. 1.3). Es lässt sich beweisen, dass Faltungen gewissen Grundgesetzmäßigkeiten folgen müssen. Deswegen sind diese auch bei allen Ordnungen, bei denen Flügelfaltung auftritt, verwirklicht. Der intuitive Schluss, dass die ähnliche Konstruktion auf einem gemeinsamen Vorfahren zurückzuführen ist, ist also nicht berechtigt. Vielmehr legen die paläontologischen Funde nahe, dass die Faltungsmechanismen innerhalb der Ordnungen unabhängig voneinander erfunden wurden. Die Längsfaltung ist älter und kommt in acht rezenten Ordnungen vor. Die später auftretende Querfaltung (Abb. 1.4) findet man in vier rezenten Ordnungen, wo sie vermutlich unabhängig voneinander entwickelt wurde. Der dritte Faltungstyp, der entsprechend einem Fächer arbeitet, kommt in sechs rezenten Ordnungen vor und ist ebenfalls auf keinen gemeinsamen Vorfahr zurückzuführen (Abb. 1.5).
Abb. 1.5 Fächerfaltung bei einem Ohrwurm 
Im nebenstehenden Bild sind auf der linken Seite Vorder- und Hinterflügel in Ruhestellung. Der Vorderflügel bedeckt den Hinterflügel größtenteils. Von diesem ragt lediglich das Gelenk in Form eines Viertelkreises mit einem zentralen weißen Fleck unter dem Vorderflügel hervor. Auf der rechten Bildseite ist der Vorderflügel nach rechts aufgeklappt (blauer Pfeil). Perspektivisch erscheint er schmäler als er ist. Der Hinterflügel ist noch völlig gefaltet. 
Im nebenstehenden Bild auf der linken Seite ist der Vorderflügel nach links geöffnet (Blauer Pfeil). Auf der rechten Seite wurde der Vorderflügel entfernt. Der Hinterflügel ist halb geöffnet. Bei größerer Auflösung ist die Vielfachfaltung wie bei einem Fächer parallel zu den Linien b und c deutlich erkennbar. Der Pfeil e zeigt auf den Punkt, an dem der geschlossene Fächer nochmals um 180° gefaltet wird. Schließlich wird der doppelte Stapel von Flügelfeldern unter das große Flügelfeld geschoben wird, das parallel zur Linie a liegt. Der Umfang an fossilen Funden reicht bei weitem noch nicht aus, die offenen Fragen bezüglich der Entwicklung des Insektenflügels mit einer abgeschlossenen und allgemein akzeptierten Theorie zu beantworten. Sie liefern jedoch Material für interessante Spekulationen. Eine wichtige Hypothese besagt, dass der widersprüchliche Selektionsdruck auf die einerseits für die Larven sperrigen Flügel andrerseits für das Flugvermögen wichtige Größe der Flügel auf zwei Arten beantwortet wurde. In einer Gruppe wurde die Flügelentwicklung auf das letzte Stadium vor der Imago reduziert, was letztendlich zur Entwicklung der vollständigen Metamorphose führte. Als Alternative wurden während der Larvalstadien die Flügel aerodynamisch nach hinten gelegt und verloren dabei ihre ursprüngliche Beweglichkeit, die sie erst bei der letzten Häutung wiedergewinnen, wie es heute bei der unvollständigen Verwandlung beobachtet werden kann. Mit den Flügeln könnte man also die Aufspaltung der Insekten in Holometabole und Hemimetabole erklären.
Eine Zusammenfassung der Fossilgeschichte der einzelnen Insektengruppen findet sich bei Rasnitsyn und Quicke.[12][3][13][14][8][15][16]
Flügelbau
Allgemein
Fluginsekten besitzen im Grundplan vier Flügel, je ein Paar am mittleren und hinteren Brustsegment. Es gibt vielfältige Abweichungen, die meist für die systematische Einteilung von Bedeutung sind und auf die weiter unten in Form einer Tabelle eingegangen wird. Allerdings können auch innerhalb einer Ordnung, manchmal sogar innerhalb einer Art oder bei den Geschlechtern große Unterschiede auftreten. Zum Beispiel kommen bei manchen Heuschreckenarten alle Übergänge von normaler Vierflüglichkeit bis zur Flügellosigkeit vor, bei den Glühwürmchen besitzen nur die Männchen Flügel usw. Die folgenden Absätze beschreiben daher nur den grundsätzlichen Flügelbau.
Die Hautflügel der Insekten bestehen aus einer doppelten Schicht Kutikula, die direkt in die Kutikula des Thorax übergeht. Diese Doppelschicht kann unabhängig voneinander in Dicke (mehr Chitin) und Härte (mehr Sklerotin) differieren. Die eigentliche Membran ist sehr dünn (nur etwa ein Mikrometer), und in einigen Bereichen, oft in Linienform wenig gehärtet, was einerseits die Beweglichkeit der Gelenkhäute bei der Einlenkung in den Thorax ermöglicht, zum anderen aerodynamisch wichtige Verformungen während des Fluges und die Faltung bei Ruhestellung erlaubt. Es existieren jedoch auch Membranbereiche in Form dünner, gehärteter Platten, die sich zwar biegen lassen, sich aber nicht verziehen können. Die Versteifung der häutigen Abschnitte und der Platten gegeneinander erfolgt durch die sklerotisierten Flügeladern. Sie sind bis zu 100 Mikrometer dick, können aber abschnittsweise in vordefinierten Richtungen und in beschränktem Maße biegbar sein. Sie sind größtenteils hohl und werden nach der Häutung mit Hämolymphe aufgepumpt, wobei sich die Flügel entfalten. Anschließend härten sie an der Luft aus und sind in der Regel durch dunklere Pigmentierung erkennbar. Die Nerven und Tracheen der Flügel, sofern vorhanden, verlaufen in den Adern. Eine Muskulatur existiert innerhalb der Hautflügel nicht.
Abb.2.1: Bewegung der Insektenflügel bei indirekter Flugmuskulatur
Schematischer Querschnitt durch ein Brustsegment mit Flügeln
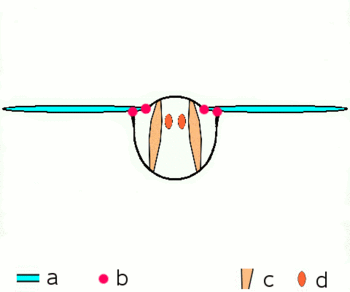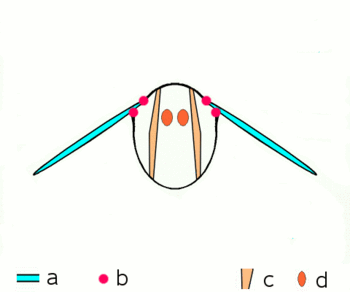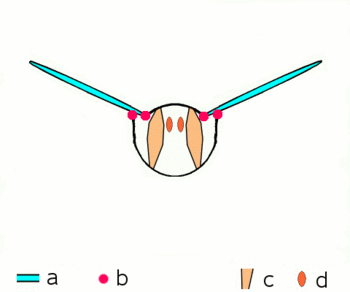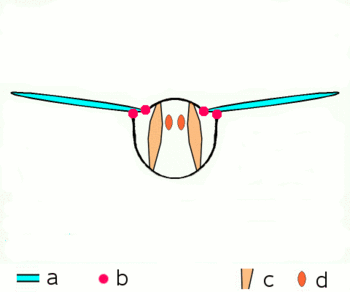
a Flügel b Gelenke
c dorsoventrale und d longitudinale Muskulatur
Die von oben nach unten verlaufende kräftige dorsoventrale Muskulatur (c) verformt mit ihrer Kontraktion den elastischen Brustring, dabei bewegen sich die Flügel nach oben. Danach entspannt sich die dorsoventrale Muskulatur, der Brustring schnellt in seine natürliche Stellung zurück, unterstützt durch die Kontraktion der Längsmuskulatur (d). Dabei bewegen sich die Flügel nach unten. Näheres zu den Gelenken siehe untenDie Verstrebungen der Flügel durch die röhrigen Adern erlauben Drehungen und Verzerrungen nur in beschränktem Maße und in bestimmten Richtungen und Winkeln. An der Flügelbasis bilden die Adern mit der Kutikula des Thorax in Form verschiedener Schultersklerite ein Gelenksystem, das die Beweglichkeit beim Flug garantiert und die Ruhestellung der Flügel ermöglicht. Die phylogenetisch älteren Flugbewegungen werden über Hebel durch abwechselnde Kontraktionen der Brustmuskulatur in vertikaler Richtung und in Richtung der Körperachse hervorgerufen (Abb. 2.1). Wenn sich die vom Rücken zum Bauch verlaufende Brustmuskulatur zusammenzieht, wird der Flügel gehoben, kontrahiert die Muskulatur, die die Brust von vorn nach hinten durchzieht, wird die vorhergegangene Abplattung der Brust wieder aufgehoben und der Flügel wird gesenkt. Weitere Muskelgruppen, die an der Unterseite der Basis der Flügel ansetzen, regeln die Flügelstellung und Flügelform während des Flügelschlags und erlauben bei den Neoptera eine Flügelbewegung in Form einer Acht.[13][14][16]
Aderung und Zellen
Ein charakteristisches Merkmal der Flügel ist ihre Aderung. Anhand dieser können Ordnungen und Familien und sogar einzelne Gattungen und Arten unterschieden werden.
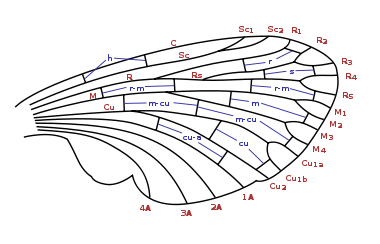 Abb.2.2: Benennung der Flügeladern der Insekten nach dem System von Comstock und Needham
Abb.2.2: Benennung der Flügeladern der Insekten nach dem System von Comstock und Needham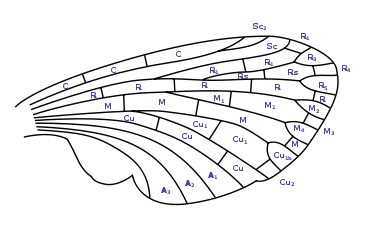 Abb.2.3: Benennung der Zellen gemäß Comstock-Needham
Abb.2.3: Benennung der Zellen gemäß Comstock-NeedhamZur Beschreibung der Aderung gibt es verschiedene Benennungssysteme. Sie bezeichnen die Adern und die dadurch entstandenen Flügelareale. Die Benennungssysteme haben dabei zwei z. T. widersprüchliche Ziele. Einerseits wäre ein Benennungssystem wünschenswert, in dem sich das ursprüngliche Adersystem widerspiegelt, so dass sich die heute auftretenden Aderungstypen als Spezialisierungen ableiten lassen. Die Ableitung der heutigen Aderungstypen von einer Urform hätte eine hohe systematische Bedeutung, ist aber bislang ungeklärt. Auf der anderen Seite stehen praktisch orientierte Benennungssysteme, mit denen die innerhalb einer systematischen Gruppe auftretende Aderungsvariationsbreite einfach und eindeutig beschrieben werden kann. In manchen Insektengruppen gibt es sogenannte Falsche Adern, die lediglich das Aussehen, nicht aber den Bau der Ader haben und deswegen in einem ursprünglichen System nicht vorgesehen werden können.
Eines der ersten und vermutlich auch das verbreitetste Modell einer systematischen Benennung der Aderung ist das 1898 entstandene Comstock-Needham System(Abb.2.2, 2.3). Es stellte ein wichtiges Mittel dar, um die Homologie der Insektenflügel zu zeigen. Das System sieht für die sechs großen Longitudinaladern im Flügel, beginnend an der Vorderkante, folgende Namen vor:
- Costa (C),
- Subcosta (Sc),
- Radius (R),
- Median (M),
- Cubitus (Cu),
- Analadern (1A, 2A,..).
Im Falle von Aufspaltungen der Adern werden ihre Namen noch mit Zahlen indiziert.
Die Unterschiede in der Bezeichnung der Adern in anderen Benennungssystemen beziehen sich häufig nur auf den hinteren Teil der Flügel. So nennt Snodgrass beispielsweise die erste Analader Postcubitus (PCu) anstatt A1 und beginnt dann ab der zweiten Analader diese als Vannaladern zu nummerieren.

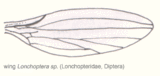
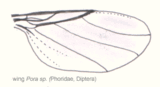
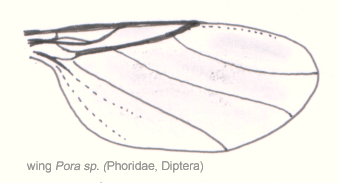
Abb. 2.4 Aderung der Trichoceridae Abb. 2.5 Aderung der Lonchopteridae Abb. 2.6 Aderung der Phoridae 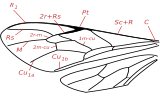
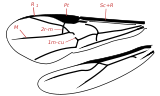
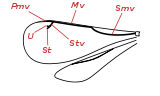
Abb. 2.7 Reduzierte Aderung bei Ichneumonidae Abb. 2.8 Stark reduzierte Aderung bei Braconidae Abb. 2.9 Extrem reduzierte Aderung bei Chalcidoidea Auch die durch die einzelnen Adern abgegrenzten Bereiche, die sogenannten Zellen, werden benannt. Eine Zelle heißt geschlossen, wenn sie auf allen Seiten durch Adern begrenzt ist, und offen, wenn eine Seite an den Flügelrand reicht. Dabei leitet sich der Name der Zelle nach Comstock-Needham von der davorliegenden Ader ab. So wird beispielsweise die Zelle zwischen Sc2 und R1 als Sc2 bezeichnet. Im Falle der Aufspaltung der Adern sehen die verschiedenen Systeme natürlich weiter differenzierte Namen vor. Für das Comstock-Needham-System ist die Benennung in Abb.2.2 dargestellt.
Um Besonderheiten im Flügelbau bestimmter Ordnungen besser beschreiben zu können, wurden weitere Benennungssysteme geschaffen. So gibt es beispielsweise für Libellen noch über fünf weitere Benennungssysteme, die parallel zu dem von Comstock-Needham genutzt werden.[13]
Bei stammesgeschichtlich moderneren Formen kann man eine zunehmende Differenzierung des Aderbaus und eine Reduzierung der Anzahl der Adern und damit der Zellen feststellen. Eine reduzierte Aderung muss jedoch nicht zwangsläufig bedeuten, dass die Insektengruppe stammesgeschichtlich jünger ist, da die Anzahl der vorhandenen Adern auch von der Größe des Insekts abhängt. Die Abbildungen 2.4, 2.5 und 2.6 zeigen die Aderung verschiedener Dipteren. Bei der knapp einen Zentimeter großen Trichoceridae sind fast alle Längsadern vorhanden, allerdings gibt es nur noch zwei Analadern und die Queradern sind stark reduziert. Die durchschnittlich kleineren Longopteriden (ca. vier Millimeter) haben weniger Adern, und bei den ein bis zwei Millimeter großen Phoridae ist die Aderung noch stärker reduziert. Das gleiche Phänomen ist bei den Hymenopterenflügeln in den Abbildungen 2.7, 2.8 und 2.9 zu beobachten. Die Ichneumoniden besitzen nur noch vier radiäre Adern, bei den kleineren Braconiden ist die Aderung deutlich reduziert und bei den Chalcidoidea, zu denen die kleinsten geflügelten Insekten gehören, ist die Aderung extrem reduziert.[17]
Felder
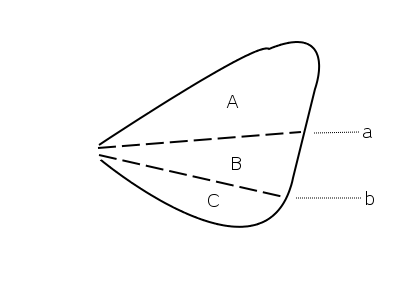
Abb. 2.10 Flügelfelder nach Wooton 1979.Der Bereich zwischen Remigium und Jugum wird bei Fehlen weiterer Faltungslinien Clavus, andernfalls Vannus genannt 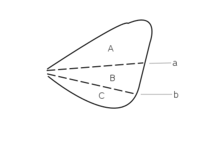
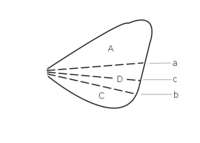
A Remigium
B Clavus
C Jugum
D Vannusa Plica clavalis
b Plica jugalis
c Plica vannalisLeider ist die Bezeichnung der verschiedenen Flügelareale ebenfalls nicht einheitlich. In der Tradition von Comstock-Needham wird der Flügel in Felder unterteilt, die durch die Längsadern begrenzt werden. Dabei benennt man die Felder im Allgemeinen nach der Ader, die das Feld nach vorn begrenzt. Das Costalfeld liegt hinter der Costa, das Subcostalfeld hinter der Subcosta, das Medialfeld hinter der Media. Das Praecostalfeld dagegen liegt vor der Costa, das Analfeld wird nach vorn nicht durch eine Ader, sondern durch die Analfalte begrenzt, und das Pterostigma wird ebenfalls häufig als Flügelfeld bezeichnet, obwohl es Teil des Costalfeldes ist. Auch hier dienen die gängigen Bezeichnungen in erster Linie dazu, innerhalb einer Insektengruppe Merkmale zu beschreiben. Im einfachsten Fall, z. B. bei den Phasmiden, unterscheidet man nur ein Costal- und ein Analfeld. Im Zusammenhang mit der Frage der Evolution der Insektenflügel sind jedoch die Faltungslinien (entlang derer der Flügel in der Ruhestellung gefaltet wird) und die Flexionslinien (entlang derer sich die Flügel beim Flug biegen) für die Begrenzung der Flügelfelder wichtig geworden (Wooton 1979, Abb. 2.10). Die wichtigsten Faltungslinien sind die Jugalfalte (Plica jugalis, hinter der dritten Analfalte) und die Clavalfalte (Plica clavalis, liegt dem Postcubitus an). Sie teilen den Flügel in drei Felder. Vor der Clavalfalte liegt das Remigium, das dem Costalfeld entspricht. Dahinter zwischen Claval- und Jugalfalte liegt der Clavus, und hinter der Jugalfalte das Jugum. Bei Hinterflügeln findet man zwischen Jugal- und Clavalfalte gelegentlich eine oder weitere Faltungslinien, die Vannalfalten. In diesem Fall wird der Bereich zwischen Claval- und Jugalfalte, in dem die Vannalfalten liegen, nicht als Clavus, sondern als Vannus bezeichnet, sodass sich die Felder Remigium, Vannus und Jugum ergeben. .[18][3]
Gelenke
An der Basis des Flügels laufen alle Längsadern zusammen und sind dort entweder direkt oder mittels Syndesen mit den Skleriten des Rückens verbunden. Die Sklerite werden auch Axillaria genannt. Für die einzelnen Sklerite haben sich folgende Namen eingebürgert. Der an die Costa anschließende Sklerit wird Humeralsklerit genannt, die danach folgenden die Pterale 1 bis 3 die in dieser Reihenfolge zu Subcosta, Radius und Anales gehören. Bei einigen Arten kommt auch noch ein mit keiner Ader verbundenes Pterale 4 vor. Diese Sklerite bilden zusammen das sekundäre Flügelgelenk. Das primäre oder pleurale Flügelgelenk bildet sich auf der Flügelunterseite durch das Pteral 2 und dem Fulcrum die über eine Membran verbunden sind. Das Pteral 3 ist dafür verantwortlich das die Flügel angelegt werden können.[18]
Flügelbau und Systematik
Schon Aristoteles benannte Gruppen von Insekten nach dem Bau ihrer Flügel. Heute werden fast alle Insektenordnungen mit der Endung -ptera (gr. πτερόν = Flügel, bei Insekten im Sinne von häutigem Flügel) gebildet. Stark vereinfachend kann man sagen, dass der Flügelbau die Insektenordnungen definiert, ihre Aderung die Familien. Bei höheren systematischen Einheiten wird häufig -oidea (εἶδος Art, -artig) angehängt, so dass sich die Endung -pteroidea ergibt. Außerdem werden auch Gruppen von Fluginsekten, bei denen kein engerer verwandtschaftlicher Zusammenhang gegeben ist oder dieser umstritten ist mit Wörtern belegt, in denen der Wortteil -ptera vorkommt. Der erste Wortteil weist auf eine Eigenheit der Flügel hin, die die Gruppe charakterisiert. Die folgende Tabelle ist nach den wissenschaftlichen Ausdrücken alphabetisch geordnet, so dass sie als Glossar benutzt werden kann.
Wissenschaftlicher Name sprachliche Wurzel Übersetzung des Wissenschaftlichen Namens Deutscher Name Anmerkungen Anisoptera ἀνισο- aniso- ungleich Ungleichflügler Großlibellen Unterordnung, im Unterschied zu den Kleinlibellen sind Hinter- und Vorderflügel verschieden groß. Bemerkung 7 Aptera
apterἀ- ohne Flügellose
flügellos---
flügellosOrdnung bei Linnaeus, heute aufgelöst, Insekten können primär oder sekundär flügellos sein, Bemerkung 1 Apterygota πτερύγιον pterygion (kleiner) Flügel
ἀ ohneFlügellose Urinsekten (plesiomorphe) Unterklasse, Zusammenfassung aller primär flügellosen Insektenordnungen, vgl. Bemerkung 1 Coleoptera κολεός koleos lederne Hülle, in die das Schwert gesteckt wurde (Schwertscheide) Umhüllt-Flügler Käfer Ordnung, Vorderflügel bilden eine schützende Hülle. Bemerkung 5 Dermaptera δέρμα derma Haut, Leder Lederflügler Ohrwürmer Ordnung, Vorderflügel verkürzt und ledrig, Bemerkung 5 Diaphanopteroidea διαφανής diaphanēs durchsichtig, durchscheinend mit durchsichtigen Flügeln --- Ausgestorbene Überordnung Dictyoptera δίκτυον diktyon Netz Fluginsekten mit netzförmiger Aderung Schaben, Termiten, Fangschrecken Ordnung, Systematik ungeklärt Diptera δύο dyo zwei Zweiflügler Fliegen, Schnaken Ordnung, nur ein häutiges Flügelpaar, Bemerkung 3 Embioptera ἐν- en- innen
βίος bios LebenInnenlebende Fluginsekten Tarsenspinner Ordnung; spinnen Röhren, in denen sie leben; nur Männchen geflügelt Endopterygota ἐντός entos innen
πτερύγιον (kleiner) FlügelInnenflügler --- Die Flügel entwickeln sich im Körperinnern;
Pterygota mit vollständiger Verwandlung;
synonym zu HolometabolaEphemeroptera ἐφήμερος ephēmeros über (einen) Tag (lang) Eintagsflügler Eintagsfliegen Ordnung, auch letztes Larvenstadium geflügelt; dieses und Adult sehr kurzlebig Exopterygota ἔξω exō außen Außen-Fluginsekten --- Flügel entwickeln sich außen am Körper;
Pterygota mit unvollständiger Verwandlung,
synonym zu HemimetabolaHemiptera ἡμι- hēmi- halb- Halbflügler Schnabelkerfe (Wanzen, Zikaden, Blattläuse, …) Bemerkung 2, Bemerkung 5 Heteroptera ἑτερο- hetero- verschieden Verschiedenflügler Wanzen Vorder und Hinterflügel sind verschieden,
Bemerkung 2, Bemerkung 5Homoptera ὅμο- homo- gleich-, ähnlich- Gleichflügler Zikaden, Blattläuse … Vorder- und Hinterflügel sind nicht wie bei Heteroptera verschieden; Bemerkung 2, Bemerkung 5 Hymenoptera ὑμένιον hymenion (dünne) Membran (Dünn)Hautflügler Hautflügler (Bienen, Ameisen, …) Ordnung, Bemerkung 4 Isoptera ἶσον īson gleich Gleichflügler Termiten Vorder- und Hinterflügel sind gleich Lepidoptera λεπίς lepis, Genitiv: λεπίδος lepidos Schuppe Schuppenflügler Schmetterlinge Ordnung, die Flügel tragen Schuppen. Bemerkung 8 Lonchopteridae λόγχη Lanze(nspitze) Lanzenfliegen Lanzenfliegen Familie der Dipteren, Flügel hat Form einer Lanzenspitze Mecoptera μῆκος mēkos Länge Langflügler Schnabelfliegen, Schnabelhafte
(Skorpionsfliegen …)Ordnung Megaloptera μεγαλο- groß- Großflügler Schlammfliegen Ordnung oder Unterordnung, Bemerkung 11 Neuroptera νεῦρον neuron Nerv, Ader Aderflügler Netzflügler Ordnung, Flügel stark netzartig geadert Neoptera νέος neos neu/jung Neuflügler --- umfasst alle derzeit lebenden Ordnungen von Fluginsekten außer Eintagsfliegen und Libellen Oligoneoptera ὀλίγον oligon wenig
νέος neos neuNeoptera mit wenig Adern --- Synonym für Holometabola, v.a. bei russischen Taxonomen beliebt. Orthoptera ὀρθο ortho- gerade, richtig Geradeflügler Heuschrecken, Grillen… Systematik umstritten, Flügel laufen gerade nach hinten Palaeodictyoptera παλαιός palaios alt
δίκτυον diktyon NetzAlt-Netzflügler --- ausgestorbene Ordnung Palaeoptera παλαιόν palaion alt Altflügler --- Systematik nicht geklärt, hauptsächlich ausgestorbene Ordnungen, rezente Ordnungen ohne Mechanismus zum Umklappen der Flügel nach hinten Paraneoptera παρα- anstatt, für
νέος neuNeoptera mit unechten Flügeln (Hemielytren, Paraelytren, …) Wanzen, Ohrwürmer … Division der Neoptera, Systematik ungeklärt Phthiraptera φθείρ phtheir Laus
ἀ ohneTierläuse Tierläuse Ordnung flügelloser Ektoparasiten, Bemerkung 10 Plecoptera πλέκειν plekein falten Faltflügler Steinfliegen Ordnung, Flügel in Ruhe um den Körper gewickelt. Polyneoptera πολύς polys viel
νέος neos neuNeoptera mit vielen Adern --- Division der Neoptera, Systematik ungeklärt Psocoptera ψώχω psocho reiben Fluginsekten, die nagen Staubläuse Ordnung, Vorder- und Hinterflügel in Ruhe und beim Flug gekoppelt, mit nagenden Mundwerkzeugen Pterostigma στίγμα stigma Fleck, Mal, Wunde Flügelfleck Flügelmal opaker oder pigmentierter Fleck nahe 1. Flügelader (Costa) bei Libellen, Staubläusen und Hautflüglern Pterothorax θώραξ thōrāx Brust „Flügelbrust“ --- Gesamtheit der Brustabschnitte, an denen Flügel sitzen, also die ganze Brust außer dem 1. Segment Pterygota πτερύγιον pterygion (kleiner) Flügel Flügel-Insekten „Fluginsekten“ Unterklasse, Gegensatz zu Apterygota, umfasst geflügelte und sekundär ungeflügelte Ordnungen, Bemerkung 1 Raphidioptera ῥαφίς Nadel Nadelflügler Kamelhalsfliegen Ordnung oder Unterordnung, Bemerkung 11 Siphonaptera σίφων siphōn Saugrohr, Rüssel
ἀ ohneRüssel-Flügellose Flöhe Ordnung, sekundär flügellos (Bemerkung 1), mit Saugrüssel Strepsiptera στρέψις strepsis Drehung Drehflügler Fächerflügler Ordnung, die Vorderflügel, die zu Halteren umgebildet sind, drehen sich beim Trocknen des Präparates, Bemerkung 3 Thysanoptera θύσανοι thysanoi Fransen Fransenflügler Fransenflügler, Thripse Ordnung, Bemerkung 6 Trichoptera τρίχωμα trichōma Haar Haarflügler Köcherfliegen Ordnung, die Flügel tragen Haare Zoraptera ζωρός zōros kräftig, stark Starkflügler Bodenläuse Ordnung, Bemerkung 9 Zygoptera ζεῦγος zeugos Paar Paarflügler Kleinlibellen Unterordnung, im Gegensatz zu den Großlibellen werden in der Ruhestellung die Vorder- und Hinterflügel paarweise zusammengelegt. Bemerkung 7 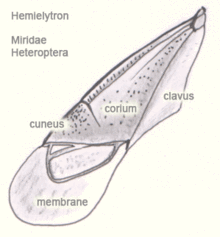 Abb. 3.1 Hemielytron
Abb. 3.1 Hemielytron Abb. 3.2 Dipterenflügel, Halteren rechts in der Ausbuchtung des Vorderflügels sichtbar
Abb. 3.2 Dipterenflügel, Halteren rechts in der Ausbuchtung des Vorderflügels sichtbar Abb. 3.3 Thrips, Flügel mit anliegenden und aufgerichteten Fransen
Abb. 3.3 Thrips, Flügel mit anliegenden und aufgerichteten Fransen Abb 3.4 Zickzackfaltung der Libellenflügel
Abb 3.4 Zickzackfaltung der Libellenflügel Abb. 3.5 Schuppen eines Schmetterlingflügels
Abb. 3.5 Schuppen eines SchmetterlingflügelsBemerkungen
- Ein Insekt heißt primär ungeflügelt, wenn seine Vorfahren sich vom Stamm der anderen Insekten abgespaltet haben, bevor die Evolution der Flügel einsetzte. Sekundär flügellos heißt dagegen ein Insekt, das geflügelte Vorfahren besitzt.[19]
- Die Heteroptera werden entweder als Ordnung gesehen, dann sind die Hemiptera eine Überordnung, oder die Hemiptera werden als Ordnung betrachtet, dann sind die Heteroptera Unterordnung. Ebenso werden die Isoptera als Ordnung, als Unterordnung der Blattodea oder (gemeinsam mit den Blattodea) als Unterordnung der Dictyoptera betrachtet. Das Wort Hemiptera bezieht sich auf die Tatsache, dass die Vorderflügel nur im hinteren Teil häutig sind. Der vordere sklerotisierte Teil ist durch eine Clavelspalte in Corium und Clavus geteilt (Abb. 3.1) Die Hinterflügel werden bei Ruhe unter den Vorderflügeln gefaltet. Bei den Homoptera sind die Flügel in Ruhe dachartig aufgestellt, bei den Heteroptera liegen sie flach auf dem Hinterleib an.[20]
- Bei den Diptera und Strepsiptera ist ein Flügelpaar zu sogenannten Schwingkölbchen (Halteren) umgebildet, kurze kleine Auswüchse, die am Ende kolbig verdickt sind. Bei den Strepsiptera sind die Vorderflügel Halteren, die Hinterflügel voll ausgebildet. Bei den Diptera sind die Hinterflügel Halteren (Abb. 3.2). Da diese nicht mehr wie Flügel aussehen, bleiben nur die zwei Vorderflügel, daher der Name Diptera, Zweiflügler. Die Halteren schwingen im Gegentakt mit den Vorderflügeln und wirken als Gleichgewichtsorgan, mit dem jede Richtungsänderung wahrgenommen werden kann. Bei manchen Gruppen dient die ausschließliche Bewegung der Halteren zum Vorwärmen der Muskulatur vor dem Start.[21][20]
- Hymenopteren besitzen im typischen Fall vier Flügel. Sie sind alle häutig. Bei einigen Familien sind auch die Vorderflügel in Ruhestellung längs gefaltet, wobei die hinteren kleiner und in der Ruhestellung unter den Vorderflügeln kaum erkennbar sind (Abb. 1.3). Mit einer Reihe kleiner Haken an der Vorderkante der Hinterflügel (Hamuli) sind diese an die Vorderflügel gekoppelt. Oft ist die Aderung stark reduziert und bildet wenige große Zellen, die für die Gruppe charakteristisch ist (Abb. 2.4 bis 2.6). An der Spitze der Vorderflügel findet man häufig ein dunkleres Pterostigma. Für die Benennung der Adern wird häufig ein eigenes System verwendet (Herbert H. Ross 1936, Weblinks)[22]
- Käfer (Coleoptera), Schaben (Blattodea), Ohrwürmer (Dermaptera, Abb. 1.5) und Schnabelkerfe (Hemiptera) sind die vier rezenten Insektenordnungen, die im Zusammenhang mit der Umbildung der Vorderflügel als Deckflügel mit Schutzfunktion die Hinterflügel auch quer falten können, um sie unter den Vorderflügeln verbergen zu können. Es handelt sich dabei nicht um getrennte Entwicklungen von Vorder- und Hinterflügeln, sondern die beiden Entwicklungen gehen Hand in Hand.
Wenn das Insekt nicht fliegt, liegen die Deckflügel teilweise oder ganz schützend über den Hinterflügeln und dem Hinterleib. Bei stark umgebildeten Vorderflügeln (Käfer, Ohrwürmer) werden diese beim Flug lediglich schräg nach oben abgespreizt und unterstützen die Stabilität und den Auftrieb. Sie führen dann keine Flugbewegungen aus und vibrieren höchstens. Nach dem Flug werden sie über den Hinterleib gelegt. Bei den Käfern berühren sich ihre Innenseiten und sind in der so genannten Naht (Sutura) parallel zur Körperlängsachse miteinander verfalzt, bei den anderen Gruppen überdecken sie sich teilweise.
Da die Hautflügel größer als die Deckflügel sind, werden sie durch Längs- und Querfaltung entlang vorgesehener Faltungslinien so zusammengelegt, dass sie unter die Vorderflügel geschoben werden können. Im einfachsten Fall (Schaben) existieren hierfür lediglich zwei zusätzliche quer verlaufende Faltungslinien, so dass wie bei Origami ein Teil des Flügels eingefaltet werden kann. Bei Käfern hat sich in der Flügelmitte ein zusätzliches Gelenksystem entwickelt. Zum Aufspannen dient die Flügelmuskulatur, die über die Form der Adern und ein entsprechendes kompliziertes System aus Schnapp- und Sperrgelenken bewirkt, dass die Flügel in aufgespannter Stellung einrasten. Diese Gelenke sind so ausgelegt, dass trotz der Kräfte, die während der Flugbewegungen auf die Gelenke wirken, die Flügel aufgespannt bleiben (Abb. 1.4). Zum Falten der Flügel nach dem Flug werden die Vorderflügel teilweise geschlossen und die Hinterflügel dann mit Hilfe der Beine und/oder des Hinterleibs darunter geschoben.[23][14][3][14] - Bei den Thysanoptera (Abb. 3.3) sind die Flügel extrem spezialisiert. Wenn Flügel vorhanden sind, sind diese 1 bis 1,2 Millimeter lang und weisen etwa 150 bis 200 der namensgebenden Fransen auf, die so dicht aneinander liegen, dass sie 20 bis 45 % der Flügelfläche bilden. Der Durchmesser der Fransen beträgt ein bis zwei Mikrometer. Vorder- und Hinterflügel der Thysanoptera sind während des Fluges durch eine Verbindung am Flügelansatz gekoppelt.[3][24]
- Libellen (Odonata): Die großen Vorder- und Hinterflügel (bis 19 Zentimeter lang und 18 Millimeter breit) sind bei den Kleinlibellen annähernd gleich groß und werden in Ruhe meist nach oben aneinandergelegt, bei den Großlibellen sind die Hinterflügel deutlich breiter als die Vorderflügel und die Flügel bleiben in Ruhe seitlich ausgebreitet. Die komplexe Aderung (Abb. 0.1) entspricht dem Benennungssystem von Comstock-Needham. Im Unterschied zu den Neoptera können die Flügel nicht nach hinten geklappt werden und besitzen keine Faltungslinien. Außerdem setzen im Gegensatz zu fast allen anderen Fluginsekten bei den Libellen die Flugmuskeln direkt an den Flügeln an. Stabilisiert werden die Flügel durch eine Reihe von Längsadern, zwischen denen die Flugfläche nicht eben, sondern abwechselnd leicht nach oben und unten geneigt aufgespannt ist (Abb. 3.4). Im Zentrum des Flügels befindet sich der Nodus (Knoten), eine querliegende Chitinverdickung, die die Adern verbindet und ihr Abknicken bei Beanspruchung während des Fluges verhindert. Am vorderen Bereich der Flügelspitze besitzen die meisten Arten ein vergrößertes und dunkel gefärbtes Flügelfeld, das als Flügelmal (Pterostigma) bezeichnet wird und durch Füllung mit Hämolymphe als Trimmtank benutzt werden kann. Die Art der Aderung wird zur systematischen Einordnung verwendet.[25]
- Lepidoptera (Schmetterlinge) Die Vorder- und Hinterflügel sind einzeln aufgehängt, werden aber im Flug in manchen Gruppen durch besondere Mechanismen durch einen Haken, das Frenulum, miteinander gekoppelt. Die Adern verlieren nach dem Austrocknen ihre Funktion. Die Flügel sind auf der Ober- und Unterseite mit Schuppen bedeckt (Abb. 3.5). Die Schuppen sind abgeflachte, artspezifische Haare, die dachziegelartig auf den Flügeln liegen und so die Flügeladern verdecken. Die einzelnen Schuppen sind immer nur einfarbig. Ihre Form variiert stark. Die häufigste Form ist die schildförmige mit drei bis fünf Spitzen und einem in einer Vertiefung verankerten schmalen Stiel am Ende. Andere sind lanzenförmig oder kreisrund. Die Schuppen sind nicht für das Fliegen notwendig. Bei den Glasflüglern (Sesiidae) sind große Bereiche der Flügel anfangs noch lose beschuppt, werden aber beim ersten Flug durch den Verlust der Schuppen durchsichtig und glasklar[26]
Für die Bestimmung wird ein eigenes Benennungssystem der Adern verwendet, das den Flügel in Regionen aufteilt. Als Beispiel werden die Flügel eines Tagfalters beschrieben: Die Regionen verlaufen vom Flügelansatz zur Spitze, wobei jeder der Vorder- und Hinterflügel in vier Regionen aufgeteilt wird. Die Adern werden auf den Vorderflügeln von 1 bis 12 nummeriert. Dabei verläuft die 1 hinten parallel zum Innenrand. Auf den Hinterflügeln befinden sich nur 9 (selten 10) durchgängige Adern. Die Flächen, die dabei von den Flügeladern begrenzt werden nennen sich Zellen bzw. Mittel- oder Diskoidalzellen. Näheres zu den Flügeln findet sich im Artikel Flügel (Schmetterling).[27] - Die Zoraptera sind eine artenarme Ordnung von sehr kleinen meist ungeflügelten Insekten meist wärmerer Gebiete. In Europa besitzen sie keine Vertreter.[3]
- Bei den Phthiraptera handelt es sich um kleine dorsoventral zusammengedrückte sekundär flügellose (Vgl. Bemerkung 1) Ektoparasiten an Vögeln und Säugern.[3]
- Die Raphidioptera besitzen vier gleiche, primitiv geaderte Flügel. Sie gehören zu den Neuroptera im weiteren Sinne und werden auch als Unterordnung der Neuroptera geführt.[3][20]
Flügelhaltung in Ruhe
Die Flügelhaltung der Insekten in Ruhe ist gewöhnlich für jede Art festgelegt und umfasst viele Möglichkeiten. Sie hat jedoch nur sehr beschränkt systematischen Wert (Vergleiche Abb. 1.2). Da die rezenten Palaeoptera die Flügel nicht nach hinten klappen können, ruhen die Großlibellen mit seitlich abgespreizten Flügeln, die Kleinlibellen und die Eintagsfliegen mit aneinanderliegend aufgestellten Flügeln. Diese Ruhestellung finden wir aber auch bei den zu den Neoptera gehörenden Tagschmetterlingen. Die Zikaden ruhen mit dachförmig aneinandergelegten Flügeln, was sie von den nahe verwandten Wanzen unterscheidet. Ebenso lassen sich die Köcherfliegen durch die dachförmig aneinander gelegten Flügel von den sehr ähnlich aussehenden Kleinschmetterlingen unterscheiden. Bei den zu den Dipteren gehörenden Schmetterlingsfliegen gibt es jedoch ebenfalls Arten, die mit dachförmig aneinandergelegten Flügeln ruhen.
Steinfliegen rollen die Flügel in Ruhestellung der Länge nach um den Hinterleib. Ähnliches finden wir auch bei einigen Käfern, die die Hinterflügel nicht falten. Dies gilt als primitives Merkmal.
Gelegentlich kann die Flügelhaltung in Ruhe für die Abtrennung zweier Arten gegeneinander hilfreich sein, z. B. bei den Schmetterlingen.[23][20][3]Ontogenese
Im Überschneidungsbereich der Fragen zur Ontogenese und zum Insektenflügel geht es darum, zu erforschen, ab welchem Zeitpunkt festgelegt ist, aus welchem Bereich des jungen Lebewesens bei normaler Entwicklung der Flügel entsteht und inwieweit dieser Bereich bei manipulierten Entwicklungen noch umprogrammierbar ist oder seinerseits umprogrammiert.
 Abb. 4.1 Libellennymphe vor dem Schlüpfen der Imago mit nach hinten gerichteten und unbeweglichen Flügelstummeln
Abb. 4.1 Libellennymphe vor dem Schlüpfen der Imago mit nach hinten gerichteten und unbeweglichen Flügelstummeln Abb. 4.2 Nymphe einer Heuschrecke
Abb. 4.2 Nymphe einer Heuschrecke Abb. 4.3 Frei gegliederte Puppe (Pupa libera) eines Käfers. Die plastisch hervortretenden Flügel sind zwischen den Beinen erkennbar
Abb. 4.3 Frei gegliederte Puppe (Pupa libera) eines Käfers. Die plastisch hervortretenden Flügel sind zwischen den Beinen erkennbar Abb. 4.4 Bedeckte Puppe (Pupa obtecta) eines Schmetterlings. Die Flügel sind nur im Umriss erkennbar
Abb. 4.4 Bedeckte Puppe (Pupa obtecta) eines Schmetterlings. Die Flügel sind nur im Umriss erkennbarAbb. 4.5. Bei der Torf-Mosaikjungfer sind die Vorgänge im Flügel nach dem Schlüpfen sichtbar: 


Nach der Häutung wird der Flügel noch weiträumig von Hämolymphe durchflossen, was durch die gelbe Färbung ersichtlich ist. Nach und nach wird der Hämolymphefluss auf Lagunen beschränkt, weshalb die Farbe ausblasst. Schließlich verkleben die beiden Hautschichten weitflächig zu der klar durchsichtigen Membran. Die Adern treten jetzt pigmentiert hervor. Die sogenannten homöotischen Gene, die direkt oder indirekt die Entwicklung steuern, sind nicht nur bei allen Insekten, sondern im gesamten Tierreich sehr ähnlich. Bei den Insekten werden bei der Eibildung und Eiablage durch die Mutter am Vorder- und Hinterende des Eies sowie dorsal wenige typische Proteine oder RNA dieser Gengruppe lokalisiert. Letztere verursachen während der ersten Zellkernteilungen die Produktion entsprechender Proteine. Die Proteine bilden durch Diffusion innerhalb des befruchteten Eies Konzentrationsgradienten, die in ihrem Zusammenspiel als Transkriptionsfaktoren eine Kaskade von Genexpressionsmustern anstoßen, die im Endeffekt zu einer Selbstregulierung der zunehmenden Differenzierung während der weiteren Entwicklung führen. Zumindest bei Drosophila ist bekannt, dass im Überlappungsgebiet des vom bereits am Ei festgelegten Vorderendes diffundierenden Bicoid-Proteins und des vom Hinterende her diffundierenden Nanos-Proteins der zukünftige Brustabschnitt und somit die Flügel entstehen.
Die Insektengruppen unterscheiden sich erheblich bezüglich des Dotterreichtums der Eier, Wanderung der Zellkerne nach den ersten Zellkernteilungen, Form und Bewegungen des Keimstreifens, Vorhandensein der Embryonalhüllen usw. Hier werden nur die grundsätzlichen Vorgänge der Entwicklung beschrieben und es wird lediglich auf die wichtigsten Unterschiede im Zusammenhang mit den Flügeln eingegangen. Insekteneier sind dotterreich, der Dotteranteil ist bei Exopterygoten durchschnittlich höher als bei Endopterygoten. Nach den ersten Zellkernteilungen ordnen sich die Zellkerne nahe der Oberfläche des Eies an und von außen beginnend bilden sich Zellwände. Dieser erste Entwicklungsabschnitt endet damit, dass das Ei von einer einlagigen Zellschicht umgeben ist. Auf ihr zeichnet sich nun der Keimstreifen ab, aus dem sich der Embryo entwickelt. Während sich der Keimstreifen streckt, wird auf ihm äußerlich eine Segmentierung und die Anlage paariger Extremitätenknospen erkennbar. Aus ihnen entstehen später Mundwerkzeuge, Beine und in manchen Gruppen Hinterleibsextremitäten; sie können aber auch zurückgebildet werden oder im embryonalen Stadium fixiert bleiben. In den folgenden Entwicklungsschritten bilden sich die Embryonalhüllen und der Keimstreifen umwächst die Dottermasse und schließt sich auf dem Rücken. Die Organsysteme bilden sich aus und die Extremitäten nehmen ihre larvale Form an. Das Ektoderm wandelt sich zur Epidermis um, die kurz vor dem Schlüpfen larvale Kutikula absondert. Aus dem Hinterleib wird Haemolymphe in Brust und Kopf gepumpt, wodurch diese anschwellen. Die Eihüllen reißen und das erste Larvenstadium schlüpft. Flügelknospen sind in diesem Stadium morphologisch nicht erkennbar. Bei der Weiterentwicklung unterscheiden sich Endo- und Ektopterygoten wesentlich.
Bei den Exopterygoten (beispielsweise Heuschrecken und Wanzen) sind in den ersten Nymphenstadien (Larvenstadien) keine Flügel zu erkennen. Früh jedoch lässt sich eine winzige Falte der Haut im oberen seitlichen Bereich des zweiten und dritten Brustsegmentes festmachen, aus der sich der Flügel entwickeln wird. Diese Falte wird mit jeder Häutung überproportional zum Körper größer und einem Flügel ähnlicher (Abb. 4.2). Mit Ausnahme der Eintagsfliegen erhalten die Flügel erst nach der letzten Häutung ihre Funktionsfähigkeit. Dabei muss betont werden, dass diese Entwicklung im dreifachen Sinne nicht kontinuierlich ist. Einmal erfolgt sie wie das Körperwachstum in Schüben nur unmittelbar nach den Häutungen, nach der Erhärtung der Cuticula gibt es bis zur nächsten Häutung keine Größenzunahme mehr. Zum anderen ist die Entwicklung in den ersten Häutungen unbedeutend und erscheint in Richtung auf die letzte(n) der gewöhnlich vier bis fünf Häutungen verschoben. Schließlich verläuft die Entwicklung nicht linear, sondern in einem Umweg entwickelt sich der Flügel anfangs in Richtung auf einen nach hinten gerichteten starren Auswuchs des Notums (Abb. 4.1, Abb. 4.2), was bereits diskutierte interessante Spekulationen über die Evolution zulässt. Vor der letzten Häutung entwickeln sich die Flügel innerhalb der Flügelknospe als vielfach gefaltete Ausstülpung mit dem oben geschilderten Bau, die breit anliegende Basis der Flügelschuppen differenziert sich zu dem schmalen Flügelgelenk. Nach der letzten Häutung strecken die Flügel sich durch Einpumpen von Haemolymphe und erhalten nach dem Aushärten ihre Funktionsfähigkeit (Abb. 4.5).
Die Endopterygoten (Käfer, Fliegen …) haben Larven, die sich deutlich vom adulten Insekt unterscheiden und keine Spuren von Flügeln erkennen lassen. Erst im Puppenstadium, das von manchen als letztes Larvenstadium betrachtet wird, in dem keine Nahrungsaufnahme stattfindet, sind die zukünftigen Flügel bereits als Ausstülpung erkennbar (offene Puppe, Abb. 4.3) oder nur in ihrer Abgrenzung angedeutet (geschlossene Puppe, Abb. 4.4). Dieses unvermittelte Auftreten beruht darauf, dass die bereits embryonal für die Flügel bestimmten Gewebebereiche während der Larvalstadien hormonell in ihrer Entwicklung gehindert wurden. Nun wird diese unterdrückte Entwicklung beschleunigt nachgeholt. In der Bandbreite der Möglichkeiten betrachten wir die zwei Extrema. Bei Arten mit großer Ähnlichkeit zwischen Larve und Adultus geschieht die Flügelbildung ähnlich wie bei den Ektopterygoten, nur dass die Flügelanlagen von außen unsichtbar im Inneren des Körpers liegen und erst im letzten Larvenstadium ausgebildet werden (z.B. bei den Schnabelfliegen). Bei Arten, deren Larven keine Ähnlichkeit zur Imago aufweisen, wie z. B. den Fliegen, erfolgt die Flügelbildung aus sog. Imaginalscheiben im Rahmen eines grundsätzlichen Umbaus des Körpers. Aus einem bereits im Embryonalstadium definierten Bereich entsteht durch Einstülpung eine Höhlung (Peripodialhöhle). Aus dem verdickten Boden dieser Höhlung stülpt sich in die Höhlung hinein die Flügelknospe. Sie ist anfangs fingerförmig und ihr Innenraum ganz von Haemolymphe durchflossen. Dann flacht sie sich ab und die Haemolympohe wird auf Lagunen beschränkt, in die Tracheen und Nerven vordringen. Schließlich verschmelzen Ober- und Unterseite der Flügelknospen großflächig, die Haemolymphe wird auf Kanäle, die zukünftigen Flügeladeren, beschränkt. Durch eine Zellverschiebung verlassen die Flügel während ihrer Entwicklung die Peripodialhöhle. Die noch nicht gestreckten und gehärteten, aber bereits voll ausgebildeten Flügel liegen am Ende des Puppenstadium dem übrigen, ebenfalls umgestalteten Körper an und werden erst nach dem Schlüpfen aus der Puppenhülle entfaltet. Im funktionsfähigen Flügel ist die doppelte Epidermislage, die die Flügelmembran abgeschieden hat, abgestorben und geschrumpft, so dass abseits der Adern die beiden Kutikulalagen direkt aufeinander liegen.
Unter der Annahme, dass die Ontogenese der Flügel die Evolution der Flügel in abgewandelter Form wiederholt, erwartet man von der Ontologie Informationen, die Rückschlüsse auf die Evolution ermöglichen. Es zeigt sich jedoch, dass auch hier die Kenntnisse, über die die sich mit diesem Fragenkomplex beschäftigende Vergleichende Entwicklungsgenetik bisher verfügt, für schlüssige Folgerungen noch zu lückenhaft und zu wenig differenziert sind. So sind bei der Fruchtfliege die Embryonalbereiche, aus denen einerseits Flügel und andrerseits Beine entstehen zum frühesten Zeitpunkt einer möglichen Identifizierung identisch, was als Beweis der Epicoxaltherorie gewertet wurde. Beim ebenfalls holometabolen Mehlkäfer jedoch liegen sie zwar nebeneinander, sind aber in ihrer Genexpression verschieden. Es ist jedoch zu erwarten, dass mit dem Anwachsen der Datenmenge zu der Entwicklung der Flügel aus ontogenetischer Sicht auch die Evolution der Insekten verständlicher wird.[3] [28][29][7]
Aufgaben des Flügels
Man kann die Funktionen der Flügel nach natürlichen Einheiten des Verhaltens wie Fortbewegung, Balz, Revierverhalten usw. ordnen oder nach bewegungstechnischen Gesichtspunkten. Die zweite Möglichkeit zieht weniger Überschneidungen nach sich und erleichtert die Erwähnung von doppelten Funktionen.
Das Fliegen
Technische Überlegungen
 Abb. 6.1 Flügelform (genauer: Tragflächen-Profil) eines typischen Flugzeugs, die für Reynoldszahlen zwischen 106 und 107 ausgelegt ist und daher für Insekten nicht geeignet ist
Abb. 6.1 Flügelform (genauer: Tragflächen-Profil) eines typischen Flugzeugs, die für Reynoldszahlen zwischen 106 und 107 ausgelegt ist und daher für Insekten nicht geeignet ist Abb. 6.2 Die Form der Vorderflügel fördert das Abgleiten von hinderlichen Gegenständen
Abb. 6.2 Die Form der Vorderflügel fördert das Abgleiten von hinderlichen GegenständenDie Hauptaufgabe der Flügel ist bei den meisten Insekten das Fliegen. In Abhängigkeit von Gewicht und Flugart des Insekts bestehen unterschiedliche Anforderungen an den Flügelbau.
Durch die im Vergleich mit Vögeln geringe Größe der Insekten müssen deren Flügel auf ganz andere Reynoldszahlbereiche hin optimiert werden (Abb. 6.1). Daher weisen die meisten Flügel nicht das typische aerodynamische Profil von Vogelflügeln auf. Vogelflügel sind für Reynoldszahlen zwischen 103 und 105 optimiert, Insektenflügel zeigen ein eher flaches Design, das für die beispielsweise bei Schmeißfliegen typischen Werte zwischen 102 und 103 besser geeignet ist. Bei Kleinstinsekten wie den weiter unten genauer beschriebenen Fransenflüglern, die mit einstelligen Reynoldszahlen zurechtkommen, ist die Flugfläche weitgehend durch Fransen ersetzt.
Flugarten
Die flache Konstruktion hat ihre Berechtigung aber nicht nur im Auftrieb. So darf ein Flügel für einen senkrechten Steigflug, wie ihn beispielsweise die Kohlschnake beherrscht, weder gewölbt sein noch ein Profil aufweisen. Andernfalls würden Kräfte in Vorwärts- oder Rückwärtsrichtung entstehen. Und auch für den Geradeausflug zeigt sich die flache Flügelform als vorteilhaft, da anders als bei den gewölbten Flügeln von Flugzeugen und Vögeln ein beidseitiges anströmen der Flügel möglich wird. Diesen Vorteil benutzen beispielsweise Insekten der Gattung Phormia, die zum Fliegen Drehschwingungen einsetzen, wofür sich die Bedeutung der Flügelseiten mehrere tausendmal pro Sekunde vertauscht. Bei der Libellengattung wird eine Spitzengeschwindigkeit von 54 km/h angegeben. Besonders erstaunlich ist dabei die nahezu zeitlose Beschleunigung.
Insekten die im sogenannten Schwirrflug auf der Stelle stehen bleiben, wie Libellen und Schwebfliegen, machen sich die Elastizität der Flügel zu Nutze. Es wird davon ausgegangen, dass die Tiere eine Wölbung der Flügel durch die Schlagart und -frequenz passiv bzw. sogar aktiv steuern können. Nachgewiesen wurde dies allerdings bislang nur für die Wanderheuschrecke. Libellen und Schwebfliegen (Abb. 0.2) sind sogar zum Rückwärtsflug fähig.
Ein weitere Flugart ist der sogenannte Gleitflug, der allerdings gewisse Ansprüche an die Flügelgröße der Insekten stellt. Längere Gleitflüge sind somit nur größeren Schmetterlings- und Libellenarten vorbehalten. Kleine Schmetterlinge können zwar auch längere Zeit gleiten sind dafür aber auf spezielle Windverhältnisse angewiesen. Der amerikanische Wanderfalter Monarch legt von Südamerika bis Nordamerika durchschnittlich 1600 km zurück, im Einzelfall wurden 2800 km Flugdistanz gemessen.[30][21]
Vorteile des Fliegens
Aus menschlicher Sicht ist das Hauptziel des Fliegens der schnelle Ortswechsel. Aus biologischer Sicht ermöglicht das Fliegen in erster Linie die Möglichkeit zur Erschließung neuer Lebensräume. Dies bedeutet aber nicht nur, dass z. B. ein Fluss überquert werden und die dortigen Nahrungsquellen erschlossen werden können oder die Blüten auf einem hohen Baum erreichbar sind. Als Folge der Beweglichkeit sind jetzt auch schneller Ortswechsel oder Wanderungen möglich. So können in kurzer zeitlicher Folge die Blüten verschiedener Bäume besucht werden oder eine Anpassung an periodische Änderungen der Umwelt ist möglich. Ebenso wichtig ist, dass jetzt verschiedene lebensnotwendige Verhaltenskomplexe örtlich getrennt werden konnten, z. B. Nahrungssuche auf dem Boden, Fortpflanzung in der Luft und Eiablage auf einem Baum.
Weiterhin ermöglicht es das Fliegen, Nahrungsquellen besser zu nutzen. Es handelt sich dabei nicht nur um das „Finden“ z. B. einer blühenden Pflanze, sondern auch um das „Überraschen“ eines möglichen Beutetiers. Parallel dazu ergibt sich die Notwendigkeit für die potentielle Beute, sich durch Flucht dem Angriff zu entziehen. So wurden nicht nur die Form der Flügel im Hinblick auf mögliche Flugmanöver hin optimiert, es war auch ein besseres Sehvermögen notwendig, bei Nachtschmetterlingen wurde ein Gegenmittel zum Ortungssystem der Fledermäuse entwickelt. Aus den zweidimensionalen Revieren werden beim fliegenden Insekt dreidimensionale Reviere mit allen Konsequenzen für die Reviermarkierung und Verteidigung.
Insgesamt ist in der Möglichkeit des Fliegens der Grund für die schnellen Radiation der Insekten zu suchen.[8]
Schutzfunktion
 Abb. 6.3 Der Käfer Dicladispa testacea
Abb. 6.3 Der Käfer Dicladispa testacea
Durch den Bau der Elytren wenig attraktiv für einen Vogelschnabel1. Mechanischer Schutz: Die Klappung der Flügel nach hinten hat zwei Konsequenzen. Von Vorteil ist, dass der Platzbedarf des Insekts geringer ist, wodurch sich neue Nahrungsquellen erschließen lassen sowie Flucht- und Versteckmöglichkeiten ergeben. Wenn sich ein Insekt jedoch in Ritzen zwängt, steigt auch die Gefahr der Beschädigung der Flügel. Es ist also vorteilhaft, wenn die Vorderflügel, die beim Klappen obenauf zu liegen kommen, gegen mechanische Beschädigungen unempfindlich sind. So haben in einigen Insektenordnungen die Vorderflügel die Aufgabe des Schutzes nicht nur der Hinterflügel sondern auch des wenig chitinisierten und verletzlichen Hinterleibs übernommen. Dieser Schutz kann auf verschiedene Art erreicht werden. Eine einfache Möglichkeit ergibt sich durch die Form, wie die Flügel in Ruhestellung dem Körper anliegen und damit z. B. das Abgleiten von Pflanzenteilen erleichtern, wenn das Insekt durchs Gras krabbelt. Ein eindrucksvolles Beispiel bietet die Schabe, deren Körperumriss bei angelegten Flügeln ideal ist, um sich in Ritzen zu zwängen (Abb. 6.2). Eine weitere Möglichkeit besteht darin, den empfindlichen Flügelhinterrand nach unten zu falten, sodass eine robustere Ader die Begrenzung der Flügelfläche im Ruhezustand bildet (Abb.1.3). Die wirksamste Möglichkeit liegt in einer stärkeren Sklerotisierung (Härtung) des Vorderflügels, die dann Deckflügel genannt werden. Dies bewirkt jedoch ein höheres Gewicht, was das Flugvermögen beeinträchtigt. Deswegen muss ein Kompromiss gefunden werden, der am besten zur Lebensweise der Art passt. Bei Arten mit schlechtem oder ohne Flugvermögen können die Vorderflügel stark sklerotisiert und miteinander verwachsen sein. Sind die Vorderflügel ganz oder teilweise deutlich verdickt und pigmentiert, zeigen aber noch eine deutliche Aderung, dann heißen sie Tegmina (Einzahl Tegmen). Sind die Deckflügel in Größe und Form dem Körperumriss des Hinterleibs angepasst und lässt sich keine Aderung mehr erkennen, dann heißen sie Elytren (Einzahl Elytron, Abb. 0.4). Ist nur ein Teil der Vorderflügel stärker ausgebildet, wird der Begriff Hemielytren (Abb 3.1) benutzt; sind die Vorderflügel nur mäßig robuster als die Hinterflügel, wird der Begriff Pseudoelytren verwendet.
Der Schutz kann darin bestehen, dass es zu keinen Verletzungen kommt, wenn die Insekten sich durchs Laub oder Gras zwängen, er kann aber auch dazu führen, dass das Insekt für einen potentiellen Fressfeind wenig attraktiv wird oder gar aus dem Speisezettel eines Insektenfressers verschwindet (Abb. 6.3).
2. Als weitere Form des Schutzes muss die Tarnung erwähnt werden, die wesentlich durch Farbe und Form der Flügeldecken verbessert werden kann (Abb. 6.4).[31][14]
Signalfunktion
 Abb. 6.4 Die Flügelhaltung kann auch zur Tarnung dienen
Abb. 6.4 Die Flügelhaltung kann auch zur Tarnung dienen Abb 6.5 Morpho peleides mit markanter blauer Färbung
Abb 6.5 Morpho peleides mit markanter blauer Färbung Abb. 6.6 Grille mit
Abb. 6.6 Grille mit Soundbeispiel?/i
Soundbeispiel?/iMit der starken Radiation der Insekten ergab sich eine breite Palette von Möglichkeiten, Individuen der eigenen Art oder anderer Arten mit den Flügeln mehr oder weniger spezielle Nachrichten zu übermitteln. Sie sind im Folgenden nach Flügelzeichnung, Flügelhaltung und Flügelbewegung gegliedert.
Färbung und Farbverteilung
Die Färbung der Flügel kann wie die des ganzen Insekts entweder auf Pigmentierung beruhen oder es treten sogenannte Strukturfarben auf. Strukturfarben ergeben sich besonders bei dünnen Schichten, wie sie eben bei Flügeln oder besonders bei den Schuppen der Schmetterlinge häufig sind (Abb. 6.5).
Farbe und Farbverteilung, die sogenannte Zeichnung, unterliegen zwar individuellen Schwankungen, sind aber in den Grundzügen artspezifisch. An der Zeichnung der Flügel erkennen viele Insekten potentielle Geschlechtspartner oder konkurrierende Artgenossen. Sie dienen als Schlüsselreiz zum Auslösen entsprechender Verhaltensketten. So kann die Intensität einer Zeichnung zum Selektionsvorteil werden. Mit der Zeichnung der Flügeldecken ahmen manche Käfer die Zeichnung des Hinterleibs von Wespen nach (Mimikry).
Flügelhaltung und -bewegung
Die Flügelhaltung signalisiert häufig die Paarungsbereitschaft. Gewöhnlich das Weibchen nimmt die Paarungsstellung ein, indem sie die Flügel so hält, dass sie dadurch die Paarung erst ermöglicht. Dies ist besonders bei Schmetterlingen leicht zu beobachten. Auch das Drohsophilaweibchen zeigt durch das Wegklappen der Flügel und Freilegen der Geschlechtsorgane seine Paarungsbereitschaft an. Es sind aber auch Drohhaltungen bekannt, die einem Angriff vorausgehen.
Die Flügelbewegung ist sowohl deutlicher sichtbar als auch plastischer als die Flügelhaltung. Sie tritt häufig im Zusammenhang mit der Balz auf. So ist das Herbeiwinken des Geschlechtspartners, das Imponieren beim Abstecken eines Reviers sowohl gegenüber dem gleichgeschlechtlichen Konkurrenten als auch dem potentiellen Paarungspartner und das Drohen bekannt.
Das Zeigen von Schreckfarben durch plötzliches Aufklappen der Flügel ist bei vielen Schmetterlingen sowie einigen Heuschrecken und Gottesanbeterinnen zu finden.
Besonders interessant ist die Flügelbewegung zur Lauterzeugung mittels Stridulation. Dabei wird eine sogenannte Schrillkante an einer Schrillleiste entlang bewegt. Die Flügel können Träger von Schrillleiste oder Schrillkante sein oder als Klangkörper die Laute verstärken. Die artspezifischen Gesänge vieler Insekten dienen zur Revierabgrenzung, dem Heranlocken potentieller Geschlechtspartner und dem Fernhalten möglicher Konkurrenten. Besonders verbreitet sind sie bei Heuschrecken und Grillen (Abb. 6.6 Grille mit Soundbeispiel), kommen aber beispielsweise auch bei Käfern vor.
Weitere Funktionen
Bei sehr vielen Insektengruppen ist es verbreitet, dass die Tiere in Ruhe ihre Flügel senkrecht zu den einfallenden Sonnenstrahlen stellen, um möglichst viel Wärme aufnehmen zu können. Damit erhöhen sie ihre Betriebstemperatur, was bei wechselwarmen Tieren bei niedrigeren Temperaturen einen Vorteil bedeutet. Ist der Körper genügend erwärmt, wird diese Stellung nicht mehr oder seltener eingenommen. Die Flügel werden also zur Wärmeregulation eingesetzt.[6]
Die Flügel werden bei Bienen als Ventilatoren benutzt, um die Luftbewegung zu erhöhen, wenn die Temperatur im Stock über 35° steigt und Wachswaben zu schmelzen drohen. Sie fächeln feuchte Luft in den Bienenstock, der durch die Verdunstungskälte abkühlt. Ebenfalls seit langem bekannt ist das Verteilen von Sexuallockstoffen beim Weibchen des Seidenspinners. Während der Jahrtausende dauernden Zucht verloren diese Schmetterlinge die Flugfähigkeit. Durch Fächelbewegungen der reduzierten Flügel wird der Duftstoff verbreitet. Er erregt die Männchen, die sich in Richtung der Duftquelle in Bewegung setzen. Manche spezialisierte Flügelschuppen, die dann meist in Feldern nebeneinander liegen und mit Haarbüscheln versehen sind, sogenannte Duftschuppen (Androkonien), produzieren Geruchsstoffe und senden sie durch Poren aus. Im Verlauf der Werbung bei der Fruchtfliege Drosophila bewirken schnelle, schwirrende Flügelbewegungen des Männchens artspezifische Töne ("Paarungsgesänge"), die das Weibchen (mit den Antennen) hört[32]. Auch der Flügelton weiblicher Stechmücken (Culicidae) dient Männchen als Signal zur Anlockung.
Einige Arten der Ölkäfer führen unter den verwachsenen Vorderflügeln eine Luftglocke, von der man vermutet, dass sie Schutz gegen die tropische Hitze und Austrocknung bietet.[33]
Erst in neuerer Zeit wurde festgestellt, dass Bienen während des Schwänzeltanzes durch Töne, die sie mittels Flügelvibration erzeugen, Informationen über die Distanz zur Futterquelle übermitteln.[3]
Bei Wasserinsekten übernehmen die Flügel teilweise Aufgaben im Zusammenhang mit dem Leben im Wasser. Bei Schwimmkäfern kann sich unter den Flügeln eine Luftglocke bilden, in die direkt Sauerstoff aus der Luft gepumpt wird oder in die im Wasser gelöster Sauerstoff diffundieren kann. Gleichzeitig beeinflusst die gespeicherte Luft den Auftrieb. Dies kann dazu eingesetzt werden, dass das Insekt durch seine Bewegung nicht dem Absinken oder Aufsteigen entgegenwirken muss. Bei der Schmetterlingsart Hydrocampa nymphaea wird die Luft in der Hülle der unter Wasser verankerten Puppe unter den Flügeln der ausschlüpfenden Schmetterlinge gehalten, sodass dieser nach dem Schlüpfen wie ein Korken an die Wasseroberfläche getrieben wird. Die parasitischen Hautflügler der Gattungen Polynema, Hydrophylax, Limnodytes, Caraphractus und Aprostocetus benutzen ihre Flügel als Ruder beim Schwimmen unter Wasser.[3][23][34]
Weblinks
 Commons: Insektenflügel – Album mit Bildern und/oder Videos und Audiodateien
Commons: Insektenflügel – Album mit Bildern und/oder Videos und Audiodateien- Bild von Deltitschala bitterfeldensis
- Weltkarten mit Fundortverteilung
- Mögliche Verwandtschaftsverhältnisse innerhalb der Palaeooptera bzw. Palaeopteroidea
- Möglichkeiten der Phylogenie der Neoptera
- Bild von Vorderbrustanhängen (Paranota), die als reduziertes 3. Flügelpaar interpretiert wurden
- Weitere Zeichnungen auf denen Insekten mit diesen Vorderbrustanhängen zu sehen sind (Jarmila Kukalová: Revisional Study of the Order Palaeodictyoptera in the Upper Carboniferous Shales of Commentry, France. Part IIII; Pyche vol. 77 March, 1970 No. I)
- Fotos fliegender Insekten
Referenzen
- ↑ Müller, F. (1873) Beiträge zur Kenntniss der Termiten. Jena. Z. Naturw. 7, 333-358;451-463.
- ↑ Grzimek’s Animal Life Encyclopedia Vol. 3 Thomson Gale ISBN 0-7876-5779-4
- ↑ a b c d e f g h i j k l Cedric Gillott: Entomology. Springer Verlag, 2005, ISBN 1-4020-3182-3 in
- ↑ Raymand C.Moore et al. MacGraw-Hill (1952) Invertebrate Fossils [1]
- ↑ Richard M Fox and Jean Walker Fox Indroduciotn to Comparative Entomology [2]
- ↑ a b Joel G. Kingsolver, M.A.R. Koehl: SELECTIVE FACTORS IN THE EVOLUTION OF INSECT WINGS. In: Annu.Rev.Entomol.. 39, 1994, S. 425-451.
- ↑ a b E.L.JOCKUSCH, K.A.OBER: Hypothesis Testing in Evolutionary Developmental Biology: A Case Study from Insect Wings. In: Journal of Heredity. 95, Nr. 5, 2004, S. 382-396.
- ↑ a b c Günter Bechly: Ur-Geziefer - Die faszinierende Evolution der Insekten. In: Stuttgarter Beiträge zur Naturkunde Serie c - Wissen für alle. 49, 2001, S. 35-38.
- ↑ Niwa, N. et al. (2010) Evolutionary origin of the insect wing via integration of two developmental modules. Evolution & Development 12(2), 168-176
- ↑ Die rasche Evolution des Insektenflügels
- ↑ Arnold H. Staniczek, Günter Bechly, Roman J. Godunko (2011): Coxoplectoptera, a new fossil order of Palaeoptera (Arthropoda: Insecta), with comments on the phylogeny of the stem group of mayflies (Ephemeroptera). Insect Systematics & Evolution, 42(2): S. 101-138, Brill, Leiden. ISSN 1399-560X (Volltext als PDF; 44 MB)
- ↑ History of insects, edited by A.P Rasnitsyn and D.L.J. Quicke in Kluwer Academic Publishers, Vorstellung mit Auszügen [3]
- ↑ a b c Klaß, Klaus-Dieter: Die Phylogenie der Dictyoptera, Cuvillier Verlag Göttingen 1995, ISBN 3-89588-363-8
- ↑ a b c d e Fabian Haas – Geometrie, Mechanik und Evolution der Flügelfaltung bei den Coleoptera, 1998, PhD Thesis, University of Jena
- ↑ Robin J. Wootton: Das Design von Insektenflügeln. In: Spektrum der Wissenschaft. Januar 1991, S. 58-65.
- ↑ a b Fabian Haas: Evidence from Folding and Functional Lines of Wings on Inter-ordinal Relationships in Pterygota. In: Arthropod Systematics & Phylogeny. 64, Nr. 2, 2006, S. 149-158.
- ↑ Paul Brohmer (Herausgeber): Fauna von Deutschland, Quelle und Meyer Heidelberg 1964
- ↑ a b Seifert, G.: Entomologisches Praktikum, Georg Thieme Verlag, Stuttgart New York 1995 ISBN 3-13-455003-2
- ↑ dtv-Atlas zur Biologie Deutscher Taschenbuchverlag München ISBN 3-423-03011-9
- ↑ a b c d Chinery, Michael Pareys Buch der Insekten Hamburg, Berlin Parey 1993 ISBN 3-490-23118-X
- ↑ a b Großes Lexikon der Tierwelt Lingen Verlag Köln
- ↑ B.N. Danforth & C.D. Michener (1988): Wing folding in the Hymenoptera. Annals of the Entomological Society of America 81(2): 342-349.
- ↑ a b c H. Freude, K. W. Harde, G. A. Lohse: Die Käfer Mitteleuropas, Bd. 9. Spektrum Akademischer Verlag in Elsevier 1966, ISBN 3-8274-0683-8
- ↑ Moritz, Gerald: Thripse. Die Neue Brehm-Bücherei Bd. 663. Westarp Wissenschaften, Hohenwarsleben 2006, ISBN 3-89432-891-6
- ↑ Jill Silsby – Dragonflies of the World [S. 180], The National History Museum, 2001, ISBN 0-565-09165-4
- ↑ Heiko Bellmann, Der Neue Kosmos Schmetterlingsführer, Schmetterlinge, Raupen und Futterpflanzen, Franckh-Kosmos Verlags-GmbH & Co, Stuttgart 2003, ISBN 3-440-09330-1
- ↑ Lionel G. Higgins und Norman D. Rilley, Die Tagfalter Europas und Nordwestafrikas (A Field Guide to the Butterflies of Britain and Europe), Verlag Paul Parey 1971, ISBN 3-490-02418-4
- ↑ W. McGinnis, M. Kuziora: Kontrollgene für den Körperbauplan. In: Spektrum der Wissenschaft. April 1994, S. 38.
- ↑ Christiane Nüsslein-Volhard: Gradienten als Organisatoren der Embryonalentwicklung. In: Spektrum der Wissenschaft. Oktober 1996, S. 38.
- ↑ Nachtigall, W.: Insektenflug, Springer-Verlag, Berlin Heidelberg New York 2003
- ↑ Peter Detzel Die Heuschrecken Baden-Württembergs Verlag Eugen Ulmer Stuttgart 1998 ISBN 3-8001-3507-8
- ↑ z.B.: Therese Ann Markow & Patrick M. O’Grady (2005): Evolutionary Genetics of Reproductive Behavior in Drosophila: Connecting the Dots. Annual Review of Genetics 39: 263–291. doi:10.1146/annurev.genet.39.073003.112454
- ↑ Bernhard Klausnitzer: Wunderwelt der Käfer. Herder Verlag Freiburg ISBN 3-451-19630-1
- ↑ Wolfgang Engelhardt: Was lebt in Tümpel, Bach und Weiher. Kosmos, Franckh’sche Verlagshandlung Stuttgart 1955


Dieser Artikel wurde am 9. Dezember 2007 in dieser Version in die Liste der lesenswerten Artikel aufgenommen. Kategorien:- Wikipedia:Lesenswert
- Entomologie
- Anatomie (Wirbellose)




































{kind=link}
{kind=link}
Wikimedia Foundation.