- Dinosaurier
-
Dinosaurier 
Plastische Lebendrekonstruktion von Tyrannosaurus rex vor dem Frankfurter Senckenbergmuseum
Zeitraum Mitteltrias bis Oberkreide
(Vögel bis Gegenwart)235 bis 65,5 (bzw. 0) Mio. Jahre Fundorte - Weltweit
Systematik Landwirbeltiere (Tetrapoda) Amnioten (Amniota) Diapsida Archosauria Ornithodira Dinosaurier Wissenschaftlicher Name Dinosauria Owen, 1842 Ordnungen - Echsenbeckensaurier (Saurischia)
- Vögel (Aves)
- Vogelbeckensaurier (Ornithischia)
Die Dinosaurier (Dinosauria, von altgriechisch δεινός deinós ‚schrecklich‘, ‚gewaltig‘ und σαῦρος sauros ‚Eidechse‘)[1] waren die Gruppe der Landwirbeltiere (Tetrapoda), die im Mesozoikum (Erdmittelalter) von der Mittleren Trias vor rund 235 Millionen Jahren bis zur Kreide-Tertiär-Grenze vor etwa 65 Millionen Jahren die festländischen Ökosysteme dominierte.
In der klassischen Systematik werden die Dinosaurier als ausgestorbene Reptiliengruppe betrachtet, aus kladistischer Sicht jedoch schließen die Dinosaurier als systematische Gruppe die Vögel, die aus einer Gruppe kleiner theropoder Dinosaurier hervorgingen, mit ein. Somit gingen nicht alle Dinosaurier während des Massenaussterbens am Ende des Mesozoikums unter, sondern überlebten als Vögel bis heute.
In anderen Wissenschaften als der Paläontologie, etwa der Vogelkunde (Ornithologie), wird weiterhin mit der zoologischen Systematik gearbeitet, nach der die Vögel eine eigenständige Klasse bilden. Das gilt ebenso für den allgemeinen Sprachgebrauch. Wenn in diesem Artikel im Folgenden von „Dinosauriern“ die Rede ist, sind damit „Nichtvogeldinosaurier“ gemeint.
Das Wissen über die Dinosaurier erhalten Paläontologen durch die Untersuchung von Fossilien, die in Form von versteinerten Knochen, Haut- und Gewebeabdrücken sowie durch Spurenfossilien überliefert sind – also durch Fußspuren, Eier, Nester, Magensteine oder versteinerten Kot. Überreste von Dinosauriern sind auf allen Kontinenten gefunden worden, einschließlich Antarktikas, da die Dinosaurier zu einer Zeit entstanden, als alles Festland im Superkontinent Pangaea vereinigt war.
In der ersten Hälfte des 20. Jahrhunderts galten Dinosaurier als wechselwarme, träge und wenig intelligente Tiere. Zahlreiche Studien seit den 1970er Jahren haben jedoch gezeigt, dass es sich um aktive Tiere mit erhöhten Stoffwechselraten und soziale Interaktionen ermöglichenden Anpassungen handelte. Dinosaurier sind zu einem Teil der weltweiten Populärkultur geworden und spielen in einigen der erfolgreichsten Bücher und Filme eine Rolle (unter anderem in Jurassic Park). Neue Entdeckungen erscheinen regelmäßig in den Medien.
Inhaltsverzeichnis
Beschreibung der Dinosaurier
 Rekonstruierte Dinosauriervergesellschaftung am „Egg Mountain“ in Montana (USA)
Rekonstruierte Dinosauriervergesellschaftung am „Egg Mountain“ in Montana (USA)
Der englische Anatom Richard Owen stellte das Taxon „Dinosauria“ im April 1842 auf. Nach der heutigen kladistischen Auffassung schließen die Dinosaurier alle Nachfahren des gemeinsamen Vorfahren von Triceratops und der Vögel ein.[2] Alternativ wurde vorgeschlagen, die Dinosaurier als alle Nachfahren des gemeinsamen Vorfahren von Megalosaurus und Iguanodon zu definieren, da dies zwei der drei Gattungen sind, die Owen bei seiner Erstbeschreibung der Dinosaurier nannte.[3] Beide Definitionen fassen dieselben Taxa zur Gruppe der Dinosaurier zusammen: Die Theropoden (zweibeinige Karnivoren), Sauropodomorphen (mehrheitlich große Pflanzenfresser mit langen Hälsen und Schwänzen), Ankylosaurier (vierbeinige Pflanzenfresser mit massiven Hautpanzern), Stegosaurier (vierbeinige, Knochenplatten tragende Pflanzenfresser), Ceratopsia (vierbeinige Pflanzenfresser mit Hörnern und Nackenschilden) und Ornithopoden (zwei- oder vierbeinige Pflanzenfresser).[4]
Die Dinosaurier waren durch eine immense Formenvielfalt gekennzeichnet. Einige waren Herbivoren (Pflanzenfresser), andere Karnivoren (Fleischfresser); einige waren quadruped (vierbeinig), andere biped (zweibeinig), und wieder andere, wie zum Beispiel Iguanodon, konnten sich sowohl biped als auch quadruped fortbewegen. Viele hatten eine Panzerung, Hörner, Knochenplatten, Schilde oder Rückensegel. Obwohl sie für eine gigantische Größe bekannt sind, variierte ihre Größe beträchtlich; so waren viele Dinosaurier nur so groß wie ein Mensch oder kleiner. Es sind keine Dinosaurier bekannt, die in aquatische Lebensräume vorgedrungen sind. Als Vorfahren der heutigen Vögel drang mindestens eine Gruppe gefiederter Dinosaurier jedoch in die Luft vor. Auch andere gefiederte Dinosaurier konnten möglicherweise fliegen.
Bis 2006 wurden 527 Dinosauriergattungen von einer geschätzten Gesamtzahl von etwa 1850 Gattungen wissenschaftlich beschrieben.[5] Eine Studie von 1995 schätzt die Gesamtanzahl auf 3400 – wovon jedoch viele nicht als Fossilien überliefert seien.[6] Im Mittel kommen derzeit pro Monat zwei neue Gattungen[7] und pro Jahr mindestens 30 neue Arten hinzu.[8]
Gruppenspezifische Merkmale
Gegenüber ihren nächsten Verwandten innerhalb der Archosaurier (Lagosuchus, Scleromochlus und den Flugsauriern) zeichnen sich die Vertreter aller Dinosauriergruppen durch eine Anzahl gemeinsamer abgeleiteter Merkmale (Synapomorphien) aus:[9]
- Schädelmerkmale: das Postfrontale fehlt; im Gaumen überlappt das Ectopterygoid das Flügelbein (Pterygoid); der Kopf des Quadratums ist in seitlicher Ansicht exponiert; Verkleinerung der Posttemporalöffnung (Fenestra posttemporalis, einer Hinterhauptsöffnung).
- Merkmale des postkranialen Skeletts (das Skelett ohne den Schädel (cranium)): die Schultergelenkpfanne ist rückwärtig orientiert; die Hand ist asymmetrisch mit verkürzten äußeren Fingern (IV und V) – bei höheren Theropoden fehlen diese völlig; das Schienbein zeigt am vorderen oberen Ende eine kammartige Erhebung (Cnemialkamm); der Astragalus (ein Fußwurzelknochen) weist einen aufwärts abgehenden Fortsatz auf; der mittlere Mittelfußknochen (an dem Zehe III ansetzt) ist S-förmig gebogen.
Während urtümliche Dinosaurier alle diese Merkmale aufweisen, kann der Knochenbau späterer Formen stark abweichen, so dass manche der aufgeführten Charakteristika nicht mehr vorhanden oder nachzuvollziehen sind.
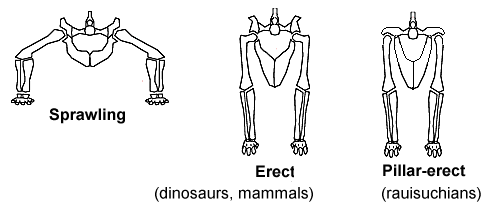
 Beckenanatomien und Gangarten verschiedener Gruppen landlebender Wirbeltiere
Beckenanatomien und Gangarten verschiedener Gruppen landlebender WirbeltiereDarüber hinaus gibt es verschiedene weitere Merkmale, die viele Dinosaurier gemeinsam haben, aber nicht als gemeinsam abgeleitete Merkmale bezeichnet werden, da sie sich gleichfalls bei einigen Nicht-Dinosauriern finden oder nicht bei allen frühen Dinosauriern auftreten. Dies sind unter anderem das verlängerte Schulterblatt (Scapula), drei oder mehr Kreuzbein-Wirbel im Bereich des Beckengürtels (drei Kreuzbeinwirbel wurden bei einigen anderen Archosauriern gefunden, jedoch nur zwei bei Herrerasaurus)[10] oder eine offene (perforierte) Hüftgelenkpfanne (geschlossen bei Saturnalia).[11]
Bei den Dinosauriern standen die Beine senkrecht unter dem Körper, ähnlich wie bei den meisten Säugetieren – aber anders als bei den meisten anderen Reptilien, deren Beine gespreizt nach außen hin abstehen (Spreizgang). Durch ihre aufrechte Haltung konnten Dinosaurier beim Bewegen leichter atmen, was wahrscheinlich Ausdauer- und Aktivitätslevel erlaubte, welche die anderer Reptilien mit gespreizten Beinen übertrafen.[12] Außerdem hat die gerade Stellung der Beine eventuell die Evolution des Gigantismus unterstützt, da so die Beine entlastet wurden.[13]
Fossilbelege
 Triceratops-Skelett im New Yorker American Museum of Natural History
Triceratops-Skelett im New Yorker American Museum of Natural HistoryDas Wissen über die Dinosaurier erhalten Paläontologen durch die Untersuchung von Fossilien, wobei Knochenfunde eine herausragende Rolle spielen – durch sie werden wichtige Daten über Verwandtschaftsbeziehungen, Anatomie und Körperbau, Biomechanik und vieles mehr gewonnen. Weitere Hinweise, besonders über das Verhalten der Dinosaurier, liefern die Spurenfossilien, etwa Zahnabdrücke an Knochen von Beutetieren, Hautabdrücke, Schwanzabdrücke, und vor allem fossile Fußspuren, die mit Abstand häufigsten Spurenfossilien. Spurenfossilien ermöglichen es, Dinosaurier aus einer anderen Perspektive zu studieren, da das Tier lebte, als die Spuren hinterlassen wurden – wobei Knochen immer von toten Tieren stammen. Weitere Informationen werden aus fossilen Eiern und Nestern, aus Koprolithen (versteinerten Kot) und Gastrolithen (Magensteine, die zur Zerkleinerung der Nahrung von einigen Dinosauriern verschluckt wurden) gewonnen.
Evolution und Systematik
Ursprung
Viele Wissenschaftler dachten lange, Dinosaurier seien eine polyphyletische Gruppe und bestünden aus miteinander nicht näher verwandten Archosauriern[14][15] – heute werden Dinosaurier als selbstständige Gruppe angesehen.[10][16]
 Lebensbild von Eoraptor lunensis, einem der ältesten bekannten Dinosaurier
Lebensbild von Eoraptor lunensis, einem der ältesten bekannten DinosaurierDie ersten Dinosaurier spalteten sich vermutlich während der mittleren Trias vor etwa 235 Millionen Jahren von anderen Archosauriern ab. Erst knapp 20 Millionen Jahre zuvor, am Ende des Perms, gab es das vermutlich größte Massenaussterben der Erdgeschichte, als 95 % aller Arten auf der Erde ausstarben.[17][18] Die 230 Millionen Jahre alte Ischigualasto-Formation in Argentinien barg die Fossilien einiger sehr alter Dinosaurier wie zum Beispiel Eoraptor, der wahrscheinlich dem gemeinsamen Vorfahr aller Dinosaurier sehr ähnlich sah[19]; somit wären die ersten Dinosaurier kleine, bipede Fleischfresser gewesen.[20] Diese Sichtweise wird ebenso durch Funde primitiver, dinosaurierähnlicher Ornithodiren bestätigt, wie Marasuchus und Lagerpeton. Diese Gattungen werden zwar außerhalb der Dinosaurier klassifiziert, waren aber wahrscheinlich mit dem gemeinsamen Vorfahren aller Dinosaurier nahe verwandt.
Als die ersten Dinosaurier erschienen, waren die terrestrischen Ökosysteme von verschiedenen Arten urtümlicher Archosaurier und Therapsiden besetzt: Aetosaurier, Cynodonten, Dicynodonten, Ornithosuchiden, Rauisuchier sowie Rhynchosaurier. Die meisten dieser Gruppen starben noch in der Trias aus; so gab es am Übergang zwischen Karnium und Norium ein Massenaussterben, bei dem die Dicynodonten und verschiedene basale Archosauromorphen wie die Prolacertiformen und Rhynchosaurier verschwanden. Darauf folgte ein weiteres Massenaussterben am Übergang zwischen Trias und Jura, bei dem die meisten anderen frühen Archosauriergruppen, wie Aetosaurier, Ornithosuchier, Phytosaurier und Rauisuchier ausstarben. Diese Verluste hinterließen eine Landfauna, die aus Crocodylomorphen, Dinosauriern, Säugetieren, Pterosauriern und Schildkröten bestand.[10]
Die frühen Dinosaurier besetzten wahrscheinlich die Nischen, die durch die ausgestorbenen Gruppen frei wurden. Früher wurde davon ausgegangen, dass die Dinosaurier die älteren Gruppen in einem langen Konkurrenzkampf zurückdrängten – dies wird heute aus mehreren Gründen als unwahrscheinlich angesehen: Die Zahl der Dinosaurier nahm nicht allmählich zu, wie es bei einem Verdrängen anderer Gruppen der Fall gewesen wäre; vielmehr machte ihre Individuenzahl im Karnium lediglich 1–2 % der Fauna aus, während sie nach dem Aussterben einiger älterer Gruppen im Norium bereits 50–90 % ausmachte. Ferner war die senkrechte Stellung der Beine, die lange als Schlüsselanpassung der Dinosaurier galt, ebenso in anderen zeitgenössischen Gruppen ausgeprägt, die nicht so erfolgreich waren (Aetosaurier, Ornithosuchier, Rauisuchier und einige Crocodylomorphen).[10]
Systematik und Phylogenese
 Zweistrahliges Becken
Zweistrahliges Becken
der Ornithischia Dreistrahliges Becken
Dreistrahliges Becken
der Saurischia Vereinfachtes Diagramm der Systematik der Dinosaurier
Vereinfachtes Diagramm der Systematik der DinosaurierDie Überordnung der Dinosaurier wird, wie die meisten heutigen Reptilien, zu den Diapsiden gezählt. Diese unterscheiden sich von den Synapsiden (aus denen die Säugetiere hervorgingen) und von den Anapsiden (die heutigen Schildkröten) durch zwei paarweise angeordnete Schädelfenster hinter den Augen. Innerhalb der Diapsiden werden sie zu den Archosauriern, den Herrscherreptilien, gezählt, die mit zwei weiteren zusätzlichen Schädelfenstern ausgestattet sind. Heutige Überbleibsel dieser Reptiliengruppe sind neben den Krokodilen die Vögel. Die Dinosaurier selber werden in zwei Ordnungen, Saurischia (auch Echsenbeckendinosaurier) und Ornithischia (auch Vogelbeckendinosaurier), aufgeteilt. Diese unterscheiden sich vorrangig an der Beckenstruktur. Die Saurischia haben die Beckenstruktur ihrer Vorfahren beibehalten und sind durch voneinander abstehende Pubis- und Ischiumknochen zu erkennen. Die Pubis- und Ischiumknochen der Ornithischia jedoch verlaufen beide parallel zueinander schräg nach hinten.
Es folgt eine vereinfachte Klassifikation von Dinosauriern auf Familienebene. Eine detailliertere Aufstellung bis auf die Ebene der Gattungen findet sich im Artikel Systematik der Dinosaurier.
- Dinosauria
- Saurischia (Echsenbeckendinosaurier: Theropoden und Sauropoden)
- Herrerasauria (frühe, bipede Fleischfresser)
- Theropoda (Bipede Dinosaurier, zum Großteil Fleischfresser)
- Coelophysoidea (Coelophysis und enge Verwandte)
- Ceratosauria (Ceratosaurus und Abelisauriden – letztere waren wichtige Predatoren der späten Kreide in den südlichen Kontinenten)
- Spinosauroidea (Fleisch und eventuell Fischfresser; einige hatten einen krokodilähnlichen Schädel und knöcherne Rückensegel)
- Carnosauria (Allosaurus und enge Verwandte, wie zum Beispiel Carcharodontosaurus)
- Coelurosauria (Gruppe verschiedenartiger Theropoden)
- Tyrannosauroidea (klein bis gigantisch, oft mit reduzierten Armen)
- Ornithomimosauria (Straußen-ähnlich, zahnlos, Fleisch- oder Pflanzenfresser)
- Therizinosauria (bipede Pflanzenfresser mit langen Armen und kleinen Köpfen)
- Oviraptorosauria (zahnlos; ihre Ernährung und Lebensgewohnheiten sind ungewiss)
- Dromaeosauridae (wie die klassischen Raptoren, zum Beispiel Velociraptor)
- Troodontidae (ähnlich wie die Dromaeosauriden, aber leichter gebaut, und möglicherweise Allesfresser)
- Aves (die Vögel, die einzigen rezenten Dinosaurier)
- Sauropodomorpha (Gruppe oft sehr langhalsiger Pflanzenfresser)
- Prosauropoda (frühe Verwandte der Sauropoden; klein bis recht groß; einige waren eventuell Allesfresser, biped und quadruped)
- Sauropoda (sehr groß, meistens über 15 Meter lang)
- Diplodocoidea (verlängerte Schädel und Schwänze; Zähne sind nach vorne gerichtet und stiftartig)
- Macronaria (diverse Gruppe teils riesiger Sauropoden)
- Brachiosauridae (sehr lange Hälse; Vorderbeine sind länger als Hinterbeine)
- Titanosauria (divers; besonders häufig in der späten Kreide der südlichen Kontinente)
- Ornithischia (Vogelbeckendinosaurier: diverse Gruppe bipeder oder quadrupeder Pflanzenfresser)
- Heterodontosauridae (kleinere Pflanzen- oder Allesfresser mit großen Eckzähnen)
- Thyreophora (Gepanzerte Dinosaurier, meistens quadruped)
- Ankylosauria (Panzerung aus Knochenplatten, einige hatten eine knöcherne Keule am Schwanzende)
- Stegosauria (quadruped, mit Knochenplatten und Stacheln)
- Ornithopoda (divers, waren gleichzeitig quadruped und biped, entwickelten Fähigkeit zu kauen, große Anzahl von Zähnen)
- Hadrosauridae (die „Entenschnäbel“)
- Pachycephalosauria („Dickkopfsaurier“, mit verdicktem Schädeldach und Kopfornamenten)
- Ceratopsia (quadrupede Dinosaurier mit Hörnern und Nackenschildern, obwohl frühe Formen nur Andeutungen dieser Merkmale hatten)
- Saurischia (Echsenbeckendinosaurier: Theropoden und Sauropoden)
Evolution, Paläobiogeographie und Palökologie
Die Evolution der Dinosaurier nach der Trias wurde durch Veränderungen der Vegetation und der Lage der Kontinente beeinflusst. In der späten Trias und im frühen Jura waren alle Kontinente zu der großen Landmasse Pangaea vereinigt, wodurch es eine weltweit einheitliche Dinosaurierfauna gab, die sich hauptsächlich aus karnivoren Coelophysoideen und herbivoren Prosauropoden zusammensetzte.[21] Nacktsamige Pflanzen, insbesondere Koniferen, verbreiteten sich während der späten Trias als mögliche Futterquelle. Prosauropoden konnten das Pflanzenmaterial nicht im Mund verarbeiten und waren auf andere Mittel zur Aufschlüsselung der Nahrung im Verdauungstrakt angewiesen.[22] Die Homogenität der Dinosaurierfaunen setzte sich bis in den mittleren und späten Jura fort: Unter den karnivoren Theropoden dominierten die Ceratosaurier, die Spinosauroideen und die Carnosaurier, während unter den Herbivoren die Stegosaurier, die Ornithischier und die Sauropoden verbreitet waren. Wichtige, gut bekannte Faunen des späten Jura schließen die der Morrison-Formation in Nordamerika und der Tendaguru Beds in Tansania mit ein. Faunen aus China zeigen jedoch bereits einige Unterschiede wie die spezialisierten Sinraptoriden unter den Karnivoren und ungewöhnliche, langhalsige Sauropoden wie Mamenchisaurus unter den Herbivoren.[21] Ankylosaurier und Ornithopoden verbreiteten sich zunehmend, die Prosauropoden jedoch starben aus. Koniferen und andere Pflanzengruppen wie Farne und Schachtelhalme waren die dominierenden Pflanzen. Anders als Prosauropoden und Sauropoden haben die Ornithischier Mechanismen entwickelt, die eine Verarbeitung von Nahrung im Mund erlaubten. So hielten backenähnliche Organe die Nahrung im Mund, und durch Kieferbewegungen konnte die Nahrung zermahlen werden.[22]
Während der frühen Kreide setzte sich das Auseinanderbrechen Pangaeas fort, wodurch sich die Dinosaurierfaunen verschiedener Kontinente mehr und mehr unterschieden. Die Ankylosaurier, Iguanodonten und Brachiosauriden verbreiteten sich über Europa, Nordamerika und Nordafrika. Später kamen besonders in Afrika Theropoden wie die großen Spinosauriden und Carcharodontosauriden hinzu; außerdem gewannen Sauropodengruppen wie die Rebbachisauriden und die Titanosaurier an Bedeutung. In Asien wurden Maniraptoren wie die Dromaeosauriden, Troodontiden und Oviraptorosaurier häufig, Ankylosaurier und frühe Ceratopsier wie Psittacosaurus wurden wichtige Herbivoren. Währenddessen wurde Australien die Heimat ursprünglicher Ankylosaurier, Hypsilophodonten und Iguanodonten.[21] Die Stegosaurier sind anscheinend in der späten Unterkreide oder der frühen Oberkreide ausgestorben. Eine große Veränderung in der Unterkreide brachte das Auftreten der Blütenpflanzen. Zur selben Zeit entwickelten verschiedene Gruppen von Herbivoren Zahnbatterien, die aus übereinander gestapelten Ersatzzähnen bestanden. Den Ceratopsiern dienten die Zahnbatterien zum Schneiden, während sie besonders bei Hadrosauriden zum Mahlen eingesetzt wurden.[22] Einige Sauropoden haben ebenfalls Zahnbatterien entwickelt, am deutlichsten sind sie bei Nigersaurus ausgeprägt.[23]
In der Oberkreide gab es drei große Dinosaurierfaunen. In Nordamerika und Asien dominierten unter den Karnivoren die Tyrannosaurier und verschiedene Typen kleinerer Maniraptoren, die Herbivoren waren überwiegend Ornithischier und setzten sich aus Hadrosauriden, Ceratopsiern, Ankylosauriern und Pachycephalosauriern zusammen. In den südlichen Kontinenten waren die Abelisauriden die vorherrschenden Predatoren und Titanosaurier die vorherrschenden Herbivoren. Die Fauna Europas schließlich setzte sich aus Dromaeosauriden, Rhabdodontiden (Iguanodontia), Nodosauriden (Ankylosauria) und Titanosauriern zusammen.[21] Blütenpflanzen breiteten sich weiter aus[22], und die ersten Gräser tauchten am Ende der Kreide auf.[24] Hadrosauriden, welche Nahrung zermahlten, und Ceratopsier, welche Nahrung lediglich abschnitten, erlangten gegen Ende der Kreide in Nordamerika und Asien eine große Häufigkeit und Vielfalt. Einige Theropoden entwickelten sich derweil zu Herbivoren oder Omnivoren (Allesfressern), wie die Therizinosaurier und die Ornithomimosaurier.[22]
Gefiederte Dinosaurier und Ursprung der Vögel
Die ersten echten Vögel tauchten im Jura auf und entwickelten sich wahrscheinlich aus den Maniraptoren,[25] einer Gruppe sehr vogelähnlicher Theropoden. Bereits 1861 wurde der berühmte Urvogel Archaeopteryx in den Solnhofener Plattenkalken entdeckt, einer einzigartigen Fossillagerstätte in der Fränkischen Alb. Dieser frühe Vogel weist ein Mosaik aus Merkmalen der Vögel und der Theropoden auf, ähnelt jedoch so sehr den Theropoden, dass mindestens ein Fossil ohne klare Federabdrücke fälschlicherweise dem Compsognathus, einem kleinen Dinosaurier, zugeschrieben wurde. Die Theorie, dass Vögel von Dinosauriern abstammen, fand jedoch erst wesentlich später allgemeine Anerkennung, nachdem sie 1970 von John Ostrom neu aufgegriffen worden war.[26] Bis heute wurden über 100 anatomische Gemeinsamkeiten zwischen theropoden Dinosauriern und Vögeln nachgewiesen[27] – Ähnlichkeiten zeigen sich besonders beim Hals, Schambein (Pubis), Handgelenk, Schultergürtel, Gabelbein und Brustbein.
Seit den 1990ern wurde eine Reihe von gefiederten Theropoden entdeckt, welche die enge Verwandtschaft zwischen Dinosauriern und Vögeln noch deutlicher erscheinen lassen. Die meisten dieser Funde stammen aus der Jehol-Gruppe aus dem Nordosten Chinas, einer Lagerstätte, die durch exzellent erhaltene Fossilien berühmt ist. Gefiederte Dinosaurier sind selten, was daran liegen könnte, dass empfindliche Strukturen wie Haut oder Federn nur selten fossilieren und deswegen in den Fossilien meistens fehlen; deswegen könnten viele andere Theropoden ebenfalls gefiedert gewesen sein. Während urtümliche, daunenartige Federn bereits bei basalen Coelurosauriern und Therizinosauroidea nachgewiesen wurden (zum Beispiel beim Compsognathiden Sinosauropteryx, beim Tyrannosauroiden Dilong und nach dem heutigen Kenntnisstand größten gefiederten Dinosaurier, dem Beipiaosaurus),[28] gehören Funde von fortgeschrittenen Konturfedern immer zur Untergruppe Maniraptora, welche die Oviraptorosaurier, die Troodontiden, die Dromaeosauriden und die Vögel umfasst.[25][29] Federn entwickelten sich ursprünglich wahrscheinlich zur Wärmeisolierung oder zur Zurschaustellung; die Funktion des Fliegens übernahmen sie erst später in der Evolution.[30][31] Bei Microraptor gui aus der Jehol-Gruppe zeigen sich gefiederte Arme und Beine – der kleine Dinosaurier könnte also eventuell mit vier Schwingen von Baum zu Baum gesegelt sein.[32] Wie sich der Vogelflug entwickelt hat, ist jedoch noch nicht geklärt.
Paläobiologie
Größe
 Die größten Dinosaurier im Größenverhältnis zum Menschen
Die größten Dinosaurier im Größenverhältnis zum MenschenObwohl Dinosaurier in der Größe stark variierten, waren Dinosaurier als Gruppe groß. Der durchschnittliche Dinosaurier war nach einer Schätzung etwa eine bis zehn Tonnen schwer,[33] während das durchschnittliche Säugetier des Känozoikums nur zwei bis fünf Kilogramm schwer wurde.[34] Einige Dinosaurier waren gar gigantisch; insbesondere die langhalsigen Sauropoden, zu denen die größten Landtiere der Erdgeschichte gehörten. Die Ursache dieses Gigantismus konnte noch nicht hinreichend geklärt werden. Für die Sauropoden nehmen einige Forscher an, dass ihre Größe Vorteile für die Verdauung brachte, da die Nahrung länger im Verdauungstrakt verweilt als bei kleineren Tieren, und so intensiver genutzt werden kann. Dies würde ein Überleben mit nährstoffarmen Pflanzen erlauben.[35]
Welcher der größte und der kleinste Dinosaurier waren, wird wahrscheinlich nie mit Sicherheit gesagt werden können. Die Überlieferung durch Fossilien ist oft sehr unvollständig, nur die wenigsten Dinosaurier sind durch vollständige Skelette bekannt – von vermutlich besonders großen Arten werden meist lediglich Knochenfragmente gefunden. Paläontologen können zwar die Form und Größe der Knochen mit denen besser bekannter Arten vergleichen, um die Größe zu schätzen – dies ist allerdings ungenau. Noch schlechter lässt sich das Gewicht der Tiere schätzen, da es unter anderem davon abhängt, wie ein Modell mit Muskeln und Sehnen versehen wird, deren Lage Paläontologen anhand von Muskelansatzstellen an den Knochen herausfinden müssen.[36]
 Brachiosaurus im Berliner Naturkundemuseum nach Abschluss der Überarbeitung 2007
Brachiosaurus im Berliner Naturkundemuseum nach Abschluss der Überarbeitung 2007Der größte und schwerste Dinosaurier, der durch gute Skelettfunde bekannt ist, ist der Sauropode Brachiosaurus (auch bekannt als Giraffatitan). Ein Skelett, das aus den Knochen verschiedener etwa gleichgroßer Individuen besteht[37], ist im Berliner Naturkundemuseum ausgestellt und hat eine Höhe von 12 Metern und eine Länge von 22,5 Meter; ein solches Tier hätte wahrscheinlich ein Gewicht von 30 bis 60 Tonnen gehabt. Der längste durch vollständige Skelette bekannte Dinosaurier ist Diplodocus, ein Skelettfund zeigt eine Länge von 27 Metern. Noch größere Sauropoden sind nur durch Skelettfragmente bekannt. Einer der größten Dinosaurier könnte Argentinosaurus gewesen sein, der manchmal auf ein Gewicht von bis zu hundert Tonnen geschätzt wird; der 33,5 Meter lange Diplodocus hallorum[35] (früher Seismosaurus) könnte zusammen mit dem 33 Meter langen Supersaurus[38] vielleicht zu den längsten Dinosauriern gehört haben.
Unter den fleischfressenden Dinosauriern gab es ebenfalls Riesen. Der größte durch fast vollständige Skelettfunde bekannte Theropode ist der etwa 12 Meter lange Tyrannosaurus rex, jedoch gibt es hier ebenso Skelettfragmente, die auf noch größere Gattungen schließen lassen. Der größte bekannte Theropode war vielleicht Spinosaurus mit einer Länge von 16 bis 18 Meter und einem Gewicht von acht Tonnen,[39] weitere sehr große Theropoden schließen Giganotosaurus, Mapusaurus und Carcharodontosaurus mit ein.
Die kleinsten Dinosaurier hatten die Größe eines Huhns – so waren die Theropoden Microraptor und Parvicursor beide weniger als 60 Zentimeter lang. Die kleineren Dinosaurier ernährten sich fast ausschließlich karnivor.
Verhalten
Interpretationen über das Verhalten der Dinosaurier basieren meistens auf der Haltung von Skelettfunden, auf Spurenfossilien wie fossilen Fußspuren, auf dem Habitat, in welchem die Tiere lebten, auf Computersimulationen der Biomechanik und auf Vergleichen mit rezenten Tieren ähnlicher ökologischer Nischen. Viele Hypothesen über das Verhalten der Dinosaurier sind lediglich spekulativ, einige finden aber Zustimmung bei den meisten Forschern.
Der Fund eines Iguanodon-Massengrabs in Bernissart (Belgien) im Jahr 1878 gab einen ersten Hinweis auf Herdenleben bei Dinosauriern.[40] Heute sind viele weitere Hinweise auf ein Herdenleben bei vielen Dinosaurierarten bekannt, so wurden neben weiteren Massengräbern auch viele parallel verlaufende Fährtenfolgen entdeckt. Hadrosauriden wanderten vermutlich in großen Herden, ähnlich wie die heutigen Springböcke oder Amerikanischen Bisons; so enthält ein Massengrab von Maiasaura aus Montana (USA) die Überreste von mindestens 10.000 Individuen.[41] Sauropodenspuren aus Oxford (England) zeigen, dass diese Sauropoden in gemischten Herden mit unterschiedlichen Arten wanderten.[42] Vielleicht bildeten Dinosaurier Herden zur Verteidigung gegen Fressfeinde, zum Schutz der Jungtiere oder für periodische Wanderungen. Einige karnivore Dinosaurier werden ebenso oft gesellig dargestellt, wobei sie als Rudeltiere zusammen selbst größere Beute erlegt haben könnten.[43][44] Jedenfalls ist Jagen im Rudel bei den nächsten lebenden Verwandten der Dinosaurier, den Vögeln und den Krokodilen, recht ungewöhnlich, und vermeintliche Rudelfunde der Theropoden Deinonychus und Allosaurus könnten die Ergebnisse von tödlichen Auseinandersetzungen zwischen fressenden Tieren sein, wie bei modernen Reptilien häufig zu beobachten.[45]
 Ein Maiasaura-Nistplatz wurde 1978 entdeckt.
Ein Maiasaura-Nistplatz wurde 1978 entdeckt.Eine Maiasaura-Nestkolonie, die Jack Horner im Jahr 1978 in Montana (USA) entdeckt hat, zeigt, dass einige Dinosaurier ihre Jungen noch lange nach dem Schlüpfen betreuten und beschützten.[46] In der Mongolei wurde 1993 das Skelett des Oviraptoriden Citipati in einer brütenden Position über seinen Eiern entdeckt; dies könnte auf isolierende Federn hinweisen, welche die Eier warm hielten.[47] Andere Funde zeigen ebenfalls elterliche Fürsorge. So wurde zum Beispiel in Liaoning (China) ein erwachsenes Exemplar des Ceratopsier Psittacosaurus zusammen mit 34 Jungtieren gefunden – die große Anzahl des Nachwuchses könnte darauf hindeuten, dass das erwachsene Tier den Nachwuchs von verschiedenen Individuen betreut hat, ähnlich wie bei heutigen Straußen.[48] Die Auca-Mahuevo-Fundstelle in Patagonien barg tausende Nester mit Eiern, die Sauropoden zugeschrieben werden und Hinweise auf große Nistkolonien dieser Dinosaurier geben, ähnlich denen der heutigen Pinguine. Sauropoden betrieben allerdings wahrscheinlich keine elterliche Fürsorge, was nicht zuletzt wegen der Größe der Elterntiere im Vergleich zu den Jungtieren angenommen wird.[49]
Die mannigfaltigen Kämme und Schilde einiger Dinosaurier, wie die der Marginocephalia, der Theropoden und der Lambeosaurinen, waren zur aktiven Verteidigung vielleicht zu zerbrechlich. Wahrscheinlicher ist, dass sie zur sexuellen Zurschaustellung dienten oder Artgenossen einschüchtern sollten – jedoch ist nur wenig über die Paarung und das Territorialverhalten der Dinosaurier bekannt. Bisswunden an den Schädeln von einigen Theropoden lassen aktive aggressive Konfrontationen zumindest bei diesen Dinosauriern vermuten.[50] Die Kommunikation der Dinosaurier untereinander bleibt ebenfalls mysteriös, ist aber ein aktives Gebiet der Forschung. Beispielsweise haben jüngere Studien gezeigt, dass die Kopfkämme der Lambeosaurinen als Resonanzverstärker für ein breites Spektrum von Rufen gedient haben könnten.[51][52]
Ein Fossil eines Troodontiden aus China zeigte, dass dieser kleine Theropode den Kopf beim Schlafen unter die Arme steckte, um ihn warm zu halten; ähnlich wie heutige Vögel.[53] Eines der für die Verhaltensforschung wertvollsten Dinosaurier-Fossilien wurde im Jahr 1971 in der Wüste Gobi entdeckt und beinhaltet einen Velociraptor, der einen Protoceratops attackiert hat; der Fund zeigt die Tiere annähernd in Lebenddarstellung.[54] Weitere Hinweise auf das Jagen lebender Beute liefert eine teilweise verheilte Schwanzverletzung eines zu den Hadrosauriern zählenden Edmontosaurus – der Schwanz wurde von einem Tyrannosaurier gebissen, das Tier überlebte aber.[55] Kannibalismus konnte bei einigen Theropoden wie Majungasaurus nachgewiesen werden; so wurden im Jahr 2003 in Madagaskar entsprechende Bissspuren gefunden.[56]
Neue Funde wie Oryctodromeus zeigen, dass einige herbivore Arten anscheinend in einem Bau unter der Erde lebten[57], während einige vogelähnliche Arten vielleicht baumbewohnend waren, wie Microraptor[32] und die rätselhaften Scansoriopterygiden.[58] Die meisten Dinosaurier bewegten sich jedoch auf dem Boden fort. Ein gutes Verständnis der Art der Fortbewegung ist ein Schlüssel für die Verhaltensforschung, und die Biomechanik hat bedeutende Fortschritte auf diesem Gebiet gebracht. So gab es Studien über die von Muskeln ausgeübten Kräfte und über das auf dem Skelett lastende Gewicht, wodurch geschätzt wurde, wie schnell Dinosaurier rennen konnten.[59] Weiter wurde untersucht, ob Diplodociden mit ihren Schwanz einen Überschallknall erzeugen konnten.[60] oder ob Sauropoden schwimmen konnten.[61] Für Theropoden wurde anhand von Kratzspuren im Sediment eines Sees nachgewiesen, dass sie schwimmen konnten.[62]
Physiologie
Seit den 1960er Jahren läuft eine energische Diskussion über die Temperatur-Regulierung der Dinosaurier. Obwohl die Theorie, dass Dinosaurier ihre Körpertemperatur überhaupt regulieren konnten, ursprünglich von Wissenschaftlern abgelehnt wurde, ist die Homoiothermie (Warmblütigkeit) der Dinosaurier die heute gängige Sichtweise, und die Debatte hat sich mehr auf die Mechanismen der Thermoregulation fokussiert.
Als die ersten Dinosaurier entdeckt wurden, glaubten die Wissenschaftler, Dinosaurier seien wechselwarme Tiere – „schreckliche Echsen“, wie ihr Name nahelegt. Man stellte sich Dinosaurier als langsame, träge Tiere vor und verglich sie mit Reptilien, die sich erst durch die Sonne aufwärmen müssen, um sich aktiv bewegen zu können. Diese Theorie einer Poikilothermie (Kaltblütigkeit) der Dinosaurier war die vorherrschende Meinung, bis Robert „Bob“ Bakker, ein früher Verfechter der Homoiothermie der Dinosaurier, eine einflussreiche Arbeit zum Thema veröffentlichte.
Hinweise auf Homoiothermie liefern Entdeckungen aus Antarktika und Australien, wo „Polar-Dinosaurier“[63] gefunden wurden, die dort einen kalten, sechsmonatigen Winter überstehen mussten. Erst kürzlich wurden Funde aus der Kreidezeit in Nordalaska gemacht, die zeigen, dass in diesen schon damals kalten Gebieten sogar dieselben Arten wie im übrigen Nordamerika lebten. Zusätzlich lässt der Skelettbau vieler Dinosaurier – insbesondere der Theropoden – auf eine hohe Aktivität schließen, die ebenfalls für eine hohe Stoffwechselrate spricht. Ebenso konnte die für homoiotherme Tiere typischen Blutgefäß-Strukturen in Dinosaurierknochen nachgewiesen werden. Eine nicht unbedeutende Anzahl von kleineren Dinosauriern verfügte außerdem über ein isolierendes Federkleid. Sauropoden zeigen jedoch weniger Anzeichen auf Homoiothermie. Es ist daher möglich, dass einige Dinosaurier warmblütig waren, andere aber nicht.[64]
Die Debatte wird dadurch verkompliziert, das Homoiothermie auf mehr als nur einen Mechanismus beruhen kann. In den meisten Diskussionen wird die Dinosaurier-Homoiothermie mit der von durchschnittlich großen Vögeln oder Säugetieren verglichen, welche Energie aufwenden, um ihre Körpertemperatur über der Umgebungstemperatur zu halten. Kleine Säugetiere und Vögel besitzen außerdem eine Isolierung in Form von Fett, Fell oder Federn, die den Wärmeverlust verringert. Jedenfalls haben große Tiere wie Elefanten ein ganz anderes Problem – wird ein Tier größer, vergrößert sich das Volumen viel schneller als die Hautfläche (Haldanes Prinzip). Ab einem gewissen Punkt übersteigt die vom Körper produzierte Wärme den Wärmeverlust über die Haut, sodass den Tieren Überhitzung droht. Besonders in Bezug auf Sauropoden wird daher die Theorie diskutiert, dass große Dinosaurier durch ihre schiere Größe wärmer als die Umgebung waren (gigantotherm), ohne dass sie spezielle Anpassungen wie Säugetiere oder Vögel besessen hätten. 2011 hatten Forscher in Science berichtet, dass die Körpertemperatur für einige große pflanzenfressende Dinosaurier auf 36 bis 38 Grad bestimmt wurde.[65]
Theropoden und wahrscheinlich ebenso Sauropoden besaßen Luftsäcke, die wie Blasebälge Luft durch die Lunge führten. Da dieses Merkmal unter heute lebenden Tieren nur bei den Vögeln bekannt ist, gilt es als weiterer Hinweis auf eine Abstammung der Vögel von den Dinosauriern. Außerdem wird es als Hinweis gedeutet, dass ebenso größere Dinosaurier wie die Sauropoden homoiotherm gewesen sein könnten, da Verdunstung in den Luftsäcken einen effektiven Kühlmechanismus darstellt.[66][67]

Computertomografische Untersuchungen von Hohlräumen in der Brustgegend des Ornithopoden Thescelosaurus zeigten im Jahr 2000 die Überreste eines komplexen, vierkammerigen Herzes.[68] In Fachkreisen besteht zwar Uneinigkeit über die Richtigkeit der Ergebnisse[69][70], das Vorkommen eines vierkammerigen Herzens sowohl bei Vögeln als auch bei Krokodilen könnte jedoch darauf hindeuten, dass Dinosaurier ebenfalls ein solches besessen haben.[71]
Im Jahre 2008 konnten Hinweise auf die Fortpflanzung der Dinosaurier gefunden werden. Bei weiblichen Vögeln wächst vor der Eiablage eine sehr calciumreiche Knochensubstanz in den Beinknochen zwischen dem harten Außenknochen und dem Knochenmark, die zur Bildung von Eierschalen benötigt wird. Eine ähnliche Knochensubstanz fand sich in den Beinknochen eines Tyrannosaurus rex, was einen ähnlichen Mechanismus vermuten und somit auf ein weibliches Tier schließen lässt.[72] Nach weiteren Forschungen wurde diese Knochensubstanz ebenso bei Allosaurus und dem Vogelbeckensaurier Tenontosaurus gefunden – da der gemeinsame Vorfahr dieser Gattungen sehr früh in der Evolution der Dinosaurier anzusiedeln ist, wird davon ausgegangen, dass alle Dinosaurier dieses Merkmal besessen haben. Da die Knochensubstanz auch bei noch nicht erwachsenen Tieren gefunden wurde, wird angenommen, dass Dinosaurier sehr schnell ihre Geschlechtsreife erreichten.[73]
Weiches Gewebe und DNA
 Scipionyx-Skelett mit Hautabdrücken in Mailand, Italien
Scipionyx-Skelett mit Hautabdrücken in Mailand, ItalienEines der besten Beispiele für Abdrücke von weichem Gewebe in einem Dinosaurierfossil wurde in Petraroia, Italien entdeckt. Der im Jahr 1998 beschriebene Fund stammt von dem kleinen, sehr jungen Coelurosaurier Scipionyx und zeigt Abdrücke verschiedener Darmabschnitte, der Leber, der Muskeln und der Luftröhre.[74]
Im Jahr 2005 stellte Dr. Mary Higby Schweitzer und ihr Team erstmals flexibles Gewebe-Material eines Dinosauriers vor, das in einem 68 Millionen Jahre alten Beinknochen eines Tyrannosaurus rex aus der Hell-Creek-Formation in Montana (USA) gefunden wurde. Nach einer Rehydrierung wurde das Material wieder elastisch, und nach einer mehrere Wochen währenden Behandlung zum Entfernen der Mineralien (Demineralisierung) konnten intakte Strukturen wie Blutgefäße, Knochenmatrix und Knochenfasern nachgewiesen werden. Genauere Untersuchungen unter dem Mikroskop zeigten darüber hinaus, dass sogar noch Mikrostrukturen auf zellulärer Ebene erhalten geblieben sind.[75] Thomas Kaye et al. stellen die Ergebnisse in einer kürzlich veröffentlichten Studie jedoch in Frage – nach diesen Forschern handelt es sich bei dem vermeintlichen Gewebematerial um bakterielle Biofilme. Die Bakterien kolonisierten einst die Hohlräume des Knochens, die zuvor von echten Zellen besetzt waren. Die von Schweitzer aufgrund ihrer Eisenhaltigkeit als Blutgefäße interpretierten Strukturen deuten die Forscher zudem als Framboide – rundliche, mikroskopisch kleine Mineralstrukturen.[76]
Die erfolgreiche Gewinnung von Dinosaurier-DNA wurde zwar in zwei Fällen gemeldet, keine dieser beiden Studien konnte jedoch bestätigt werden.[77] Jedenfalls wurde bereits das theoretische Peptid Rhodopsin eines Dinosauriers kloniert, wobei die Gene der heute lebenden Verwandten (Krokodile und Vögel) der Dinosaurier als Grundlage für die phylogenetische Ableitung dieses Peptids dienten; das in Zellkultur exprimierte und aufgereinigte Rhodopsin zeigte sich aktiv in funktionellen Tests.[78] Des Weiteren wurden verschiedene Proteine in Dinosaurierfossilien entdeckt[79], inklusive dem roten Blutfarbstoff Hämoglobin.[80] Die Möglichkeit eines Klonens von Dinosauriern, wie es in dem berühmten Roman „DinoPark“ (1993 verfilmt als „Jurassic Park“) stattfindet, ist selbst in der Zukunft eher unwahrscheinlich.
Aussterben
 Der Chicxulub-Krater nach Messungen der Schwereanomalie
Der Chicxulub-Krater nach Messungen der Schwereanomalie Künstlerische Darstellung des Dekkan-Trapp-Vulkanismus (Quelle: NSF)
Künstlerische Darstellung des Dekkan-Trapp-Vulkanismus (Quelle: NSF)In den letzten 550 Millionen Jahren gab es fünf große Massenaussterben, bei denen mindestens 40 % aller Gattungen ausstarben. Am berühmtesten ist das Kreide-Tertiär-Massenaussterben vor etwa 65 Millionen Jahren, bei dem schätzungsweise 50 % der Gattungen und 20 % der Familien verschwanden, darunter alle Nicht-Vogel-Dinosaurier. Es wurden die verschiedensten Hypothesen aufgestellt, um die Ursachen dieses Massensterbens zu klären. Die meisten aktuellen Theorien sehen die Ursache in einem Meteoriteneinschlag oder einem gesteigerten Vulkanismus, einige schließen beide Ereignisse mit ein. Das rapide Absinken des Meeresspiegels könnte gleichfalls zum Massensterben beigetragen haben, da die großen Flachmeere verschwanden und sich Landbrücken bildeten.[81]
In der Kreide-Tertiär-Grenze, einer dünnen, dunklen Sedimentschicht aus der Zeit des Massensterbens, fand Luis Alvarez eine Anreicherung des sonst in der Erdkruste seltenen Schwermetalls Iridium (siehe Iridium-Anomalie). Alvarez postulierte 1980, dass das Iridium durch einen Meteoriten auf die Erde gelangt sein könnte, welcher das Massensterben auslöste. Die Theorie wurde 1990 durch den Fund des 170 km breiten Chicxulub-Kraters am Rande der Yukatan-Halbinsel im Golf von Mexiko untermauert, der durch einen etwa 10 km großen Meteoriten entstanden ist. Die möglichen Folgen für die Ökosysteme sind schwer zu ermitteln. Forscher diskutieren beispielsweise mögliche starke Temperaturschwankungen, ebenso könnte eine Staubwolke das Sonnenlicht für einige Jahre um 10–20 % reduziert haben, was die Photosynthese der Pflanzen reduziert hätte.[82] Weitere Folgen wären wahrscheinlich Tsunamis und saurer Regen.
Ein Indiz für die Vulkanismustheorie ist der gewaltige Dekkan-Trapp-Vulkanismus der indischen Dekkan-Hochebene, der mindestens zwei Millionen Kubikkilometer Basalt (Flutbasalt, Trapp) förderte. Neben einer verminderten Sonneneinstrahlung (Insolation) könnten große Mengen Kohlenstoffdioxid in die Atmosphäre gelangt sein, was einen Treibhauseffekt zur Folge gehabt hätte.[81]
Die Dauer des Aussterbeprozesses ist umstritten. Einige Dinosaurierknochen wurden aus 64,5 Millionen Jahren alten känozoischen Schichten gefunden, was von einigen Forschern als Hinweis auf ein langsames Aussterben gedeutet wurde.[83] Andere Wissenschaftler widersprachen jedoch dieser These mit der Begründung, die Knochen seien mit der Zeit aus älteren Ablagerungen erodiert und in den känozoischen Schichten neu eingelagert worden.[84]
Eine neue Analyse vom Juli 2008 kam zu dem Ergebnis, dass die Artenvielfalt der Dinosaurier bereits seit der mittleren Kreide beständig abnahm, und widerspricht damit früheren Studien, die das Gegenteil behaupten.[85] Als die Blütenpflanzen in der mittleren Kreide einen bedeutenden Teil der Flora zu bilden begannen, stieg die Vielfalt von herbivoren Insekten und vielen anderen Gruppen, die sich an die neue Nahrungsquelle anpassten. Die Forscher vermuten, dass diese Expansion der Artenvielfalt eine Schlüsselrolle in der Entwicklung hin zu den heutigen Ökosystemen spielte. Dinosaurier jedoch blieben von dieser Expansion ausgeschlossen, was ihren Untergang einleitete, so die Forscher.[86]
Dinosaurier und Menschen
 Briefmarkenblock „Dinosaurier“ der Deutschen Post vom 4. August 2008, Serie „Für die Jugend“. Plateosaurus, Tyrannosaurus, Triceratops, Diplodocus
Briefmarkenblock „Dinosaurier“ der Deutschen Post vom 4. August 2008, Serie „Für die Jugend“. Plateosaurus, Tyrannosaurus, Triceratops, DiplodocusKeine andere Gruppe ausgestorbener Tiere hat eine derart wichtige kulturelle Bedeutung wie die Dinosaurier. Seit die Dinosaurier im 19. Jahrhundert erstmals ins Licht der Öffentlichkeit rückten, erfreuen sie sich weltweit großen Interesses und so großer Beliebtheit, dass von einer bis heute anhaltenden „Dinomanie“ gesprochen werden kann.
Fast so bekannt wie die Dinosaurier selbst ist die Tatsache, dass sie innerhalb kürzester Zeit ausgestorben sind. Daher wird der Name Dinosaurier oft als Metapher für Denk- und Handelsweisen oder für Dinge benutzt, die als rückwärtsgewandt und nicht mehr zeitgemäß empfunden werden, wie etwa beim Dinosaurier des Jahres.
Entdeckungsgeschichte
Die ersten Dinosaurierfossilien sind schon vor hunderten, wahrscheinlich tausenden von Jahren gefunden worden, wobei ihre wahre Natur nicht erkannt wurde. Im alten China in der östlichen Jin-Dynastie berichtete Chang Qu (常璩) in seinem Buch Huayang guozhi (华阳国志) über die Entdeckung von „Drachenknochen“ aus der Provinz Sichuan, die vielleicht von Dinosauriern stammten.[87] Dorfbewohner in Zentralchina haben derartige Drachenknochen seit Jahrzehnten ausgegraben, um aus ihnen traditionelle Medizin herzustellen.[88] Die antiken Griechen und Römer fanden ebenso entsprechende Fossilien, die Stoff für ihre Legenden und Sagen boten.
 Ausgrabungstätigkeiten in der spanischen Fossilfundstelle Lo Hueco 2007
Ausgrabungstätigkeiten in der spanischen Fossilfundstelle Lo Hueco 2007Im Jahr 1677 fertigte Robert Plot die erste formelle Beschreibung eines Dinosaurierfossils an, das in der Nähe von Cornwell bei Oxfordshire (England) entdeckt und heute Megalosaurus zugeschrieben wird. Obwohl er den großen Knochen zuerst einem Elefanten zuordnete, der mit den Römern nach Britannien gekommen war, erkannte Plot später eine scheinbare Ähnlichkeit mit Menschenknochen und schrieb ihn einem Riesen der biblischen Vorsintflut zu. Einer der ersten, der über diese Gruppe urzeitlicher Riesenreptilien wusste und danach forschte, war der englische Arzt Gideon Mantell. Bereits im Jahre 1822 fand er den ersten fossilen Zahn, den er einige Jahre später und nach weiteren Funden Iguanodon nannte.[89] 1824 beschrieb der Amateurpaläontologe William Buckland mit Megalosaurus erstmals einen Dinosaurier in einem wissenschaftlichen Journal. Den Begriff Dinosauria prägte jedoch ein anderer, der englische Anatom Richard Owen. Im Jahr 1842 fasste er Megalosaurus und Iguanodon mit einer weiteren Gattung, Hylaeosaurus, zu einer Gruppe zusammen, die er Dinosauria nannte.[90]
 Othniel Charles Marsh
Othniel Charles Marsh Edward Drinker Cope
Edward Drinker CopeIm Jahr 1858 wurde dann das erste fast vollständige Dinosaurierskelett in Nordamerika entdeckt. Dieser als Hadrosaurus foulkii beschriebene Fund aus Haddonfield (New Jersey) zeigte, dass dieser Dinosaurier wohl zweibeinig lief, und revolutionierte damit das öffentliche Bild der Dinosaurier – zuvor stellte man sich Dinosaurier wie Megalosaurus als riesige, auf vier Beinen laufende, waranähnliche Wesen vor. Diese Entdeckung löste eine wahre Dinomanie (Dinosaurier-Enthusiasmus) in den USA aus.
In den Folgejahren begann eine Feindschaft zwischen zwei berühmten Dinosaurierforschern, Edward Drinker Cope und Othniel Charles Marsh, die in den berühmten „Knochenkriegen“ eskalierte. Vielleicht begann der Streit, als Cope scharfe Kritik seitens Marsh erhielt, als er den Schädel des neu entdeckten, seltsamen Meeresreptils Elasmosaurus am falschen Ende des Körpers platzierte. Dies war der Beginn von Missgunst und Eifersucht zwischen den beiden Forschern und ihr Streit endete erst 30 Jahre später im Jahr 1897 nach dem Tod Copes. Jeder der beiden Kontrahenten versuchte mit seinem Team, immer mehr Dinosaurierknochen zu finden als der andere – mit allen Mitteln. Sie zerstörten sich gegenseitig viele Knochenfunde, weitere Knochen fielen dem Dynamit zum Opfer, mit dem damals Knochen freigesprengt wurden. Das Resultat der Rivalität waren 142 neu entdeckte Dinosaurierspezies, wozu Marsh 86 Arten und Cope 56 Arten beitrug.[91]
Seitdem werden auf der ganzen Welt Dinosaurierfossilien gefunden: So startete das Berliner Museum für Naturkunde eine große Expedition unter der Leitung von Werner Janensch nach Deutsch-Ostafrika, dem heutigen Tansania, die einzigartige Funde wie Brachiosaurus oder Kentrosaurus zutage förderte. Weitere wichtige Entdeckungen wurden unter anderem in Südamerika, Madagaskar, Indien, der Mongolei, China und ebenso in Deutschland gemacht.
Obwohl Dinosaurier anfangs als lebhafte, agile Tiere galten, wurde dieses Bild durch die Entdeckungen von Marsh und Cope verändert; Dinosaurier wurden zunehmend als dumme, langsame und unbeholfene Kreaturen betrachtet. Einen Sauropoden beschrieb Marsh aufgrund seines im Vergleich zur Körpergröße lächerlich klein erscheinenden Kopfes sogar als Morosaurus („dumme Echse“), später hatte sich jedoch Copes Bezeichnung Camarasaurus durchgesetzt. Erst seit den 1970er Jahren näherte sich die wissenschaftliche Meinung wieder dem ursprünglichen Bild von lebhaften, aktiven Tieren an, nachdem John Ostrom Deinonychus beschrieben hatte und die Idee von einer Warmblütigkeit der Dinosaurier aufkam. Diese Entwicklung löste die Dinosaurier-Renaissance aus, eine bemerkenswerte Zunahme von Aktivitäten innerhalb der Dinosaurierforschung, die bis heute andauert. Derzeit stammen die meisten neuen Funde unter anderem aus China und Südamerika, insbesondere Argentinien.
Dinosaurier in der Kultur
 Skulptur eines Megalosaurus von 1854 im Crystal Palace Park, London
Skulptur eines Megalosaurus von 1854 im Crystal Palace Park, LondonBreites öffentliches Interesse an Dinosauriern erregte erstmals der Crystal Palace Park in London, wo im Jahr 1853 eine Urlandschaft mit verschiedenen, lebensgroßen Modellen ausgestorbener Tiere modelliert wurde, die noch heute zu bestaunen ist. Der Bildhauer Benjamin Waterhouse Hawkins fertigte, unter Beratung von Owen, vier Dinosauriermodelle an. Die Popularität der Dinosaurier im Crystal Palace Park wurde so groß, dass Hawkins für ein ähnliches Projekt im Central Park in New York engagiert wurde. Eine neue Verwaltung des Parks ließ diesen Plan jedoch fallen, und die halbfertigen Modelle wurden zerstört.[92]
Seit Anfang des 19. Jahrhunderts zogen ausgestellte Skelette urzeitlicher Säugetiere eine Vielzahl von Besuchern an. Mit Reiseausstellungen, die durch die USA und Europa zogen, konnte viel Geld eingenommen werden. Daher sollten schon bald nach der Entdeckung vollständiger Dinosaurierskelette diese ausgestellt werden. Im Jahr 1868 wurde Hawkins beauftragt, erstmals Dinosaurier-Skelette (Hadrosaurus und Dryptosaurus) zu montieren und der Öffentlichkeit zu präsentieren. In einem New Yorker Museum gab es einen regelrechten Besucheransturm. In den folgenden Jahrzehnten wurden durch diese neuen Attraktionen in den Museen viele Skelette anderer urtümlicher Tiere in die Museumskeller verbannt, und in Deutschland konnten im Jahr 2006 allenfalls noch Relikte der Ägypter mit der Popularität von Dinosaurierskeletten konkurrieren.[93]
Von Beginn des 20. Jahrhunderts an wurde das Motiv Dinosaurier in Literatur und Film wirtschaftlich immer bedeutender und ertragreicher. Eine der ersten, und eine der berühmtesten Fantasy-Geschichten ist Arthur Conan Doyles „Die vergessene Welt“ („The Lost World“, 1912), die ab 1925 vielfach verfilmt wurde. Wie diese handeln viele andere dieser Geschichten von der Entdeckung eines bis dahin isolierten Gebietes, zum Beispiel im Regenwald oder auf einer Insel, wo Dinosaurier bis in unsere Zeit überlebt haben.
In der auch in Deutschland sehr erfolgreichen Zeichentrickserie Familie Feuerstein (1960–1966) gehörten Dinosaurier zum regelmäßigen Inventar. Später erschienen eine Reihe von Hollywood-Action-Filmen, in denen die Dinosaurier jedoch, mit Ausnahme von „Die vergessene Welt“ und Jurassic Park (1993), übertrieben und nicht den wissenschaftlichen Erkenntnissen entsprechend dargestellt wurden. So wurden Dinosaurier meist als menschenfressende Ungetüme, oft mit Höhlenmenschen zusammenlebend, dargestellt, was zeitlich unmöglich ist. Im Jahr 1954 zeigte der preisgekrönte tschechoslowakische Fantasy-Film „Reise in die Urzeit“, in dem Kinder auf einem Fluss immer tiefer in die Vergangenheit reisen, erstmals die Urzeittiere in den richtigen Zeitaltern.
Neueste Dinosaurierfilme sind oft Dokumentarfilme, zum Beispiel die mit großem Aufwand produzierte BBC-Serie Dinosaurier – Im Reich der Giganten (englischer Titel: “Walking with Dinosaurs”), in der versucht wird, Dinosaurier in ihrem Lebensraum darzustellen. Ironischerweise wird es erst durch die moderne und oft aufwändige dreidimensionale Bewegtgrafik (Computer Generated Imagery) glaubhaft möglich, diese uralten und oft gigantischen Kreaturen lebensecht zu bebildern. Dennoch hat im wissenschaftlichen Bereich die handwerkliche, oftmals kunstfertige und ästhetische Illustration, im englischen Sprachraum diese Disziplin manchmal als Paleoart[94] bezeichnet, ihre Bedeutung nicht verloren. Selbst in großen Kinoproduktionen wird heute immer mit Storyboards gearbeitet. Nach dem immensen wirtschaftlichen Erfolg des Films Jurassic Park, der in der Internet Movie Database 2007 als weltweit neunterfolgreichster Film geführt wird, erobert der Darsteller Dinosaurier ebenso die Welt des Computerspiels.
Die Leser und Betrachter dieser in Mode gekommenen Werke suchen wohl am ehesten nach Abenteuer und unentdeckten Welten (Realitätsflucht), Gefahr (Horror, Angst und Unheimliches), Zerstreuung (Unterhaltung) und Information und Bildung. Diese Kreaturen waren tatsächliche, real existierende „Monster“, die wirklich auf Erden wandelten. Außerdem könnte sich im Dinosaurier eine Abkehr von der Industrialisierung, eine Gegenmoderne manifestiert haben, die Sehnsucht der Menschen nach der verlorenen Natur.[95]
Siehe auch
 Portal:Dinosaurier und Verwandte – Übersicht zu Wikipedia-Inhalten zum Thema Dinosaurier und Verwandte
Portal:Dinosaurier und Verwandte – Übersicht zu Wikipedia-Inhalten zum Thema Dinosaurier und VerwandteLiteratur
- Kevin Padian und Philip Currie: Encyclopedia of Dinosaurs, 1997, Academic Press, ISBN 0-12-226810-5
- Gregory Paul: The Scientific American Book of Dinosaurs, 2000, St. Martin’s Press, ISBN 0-312-26226-4
- Gregory Paul: Dinosaurs of the Air: The Evolution and Loss of flight in Dinosaurs and Birds, 2002, The Johns Hopkins University Press, Baltimore, ISBN 0-8018-6763-0
- David Weishampel, Peter Dodson und Halszka Osmólska: The Dinosauria, 2. Ausg., University of California Press, ISBN 0-520-24209-2
- David Fastovsky und David Weishampel: The Evolution and Extinction of the Dinosaurs, 2005, 2. Ausg., Cambridge University Press, Cambridge, ISBN 0-521-81172-4
- James Farlow und Brett-Surman: The Complete Dinosaur, 1997. Indiana University Press, ISBN 0-253-21313-4
- W. J. T. Mitchell: The Last Dinosaur Book: The Life and Times of a Cultural Icon, 1998, The University of Chicago Press, Chicago, ISBN 0-226-53204-6 (Auszüge aus dem Buch)
- J. L. Sanz: Starring T. rex!: dinosaur mythology and popular culture, 2002, Indiana University Press, Bloomington, übersetzt von Philip Mason, ISBN 0-253-34153-1
Weblinks
 Commons: Dinosaurier – Album mit Bildern und/oder Videos und Audiodateien
Commons: Dinosaurier – Album mit Bildern und/oder Videos und Audiodateien Wiktionary: Dinosaurier – Bedeutungserklärungen, Wortherkunft, Synonyme, Übersetzungen
Wiktionary: Dinosaurier – Bedeutungserklärungen, Wortherkunft, Synonyme, Übersetzungen Wikiquote: Dinosaurier – Zitate
Wikiquote: Dinosaurier – Zitate- Auf Deutsch
- Zeitreise ins Erdmittelalter – Die Paläobiologie der Dinosaurier. Von Martin Sander. In: Gerold Wefer (Herausgeber): expedition Erde. 2., überarbeitete und erweiterte Auflage, 2006 (PDF-Datei; 1,79 MB)
- Dossier Dinosaurier von scinexx. Stand 15. August 2001
- Dinosaurier-Interesse (Privates Projekt mit einführenden Informationen über Dinosaurier und andere Tiere des Mesozoikums für Erwachsene und Kinder)
- Jurassic Harz (Privates Projekt zu den Dinosaurierfunden im Harz)
- Dinomedia (Privates Projekt zu Dinosauriern in Literatur, Film und anderen Medien)
- Auf Englisch
- Dinobase (Populärwissenschaftliches Projekt der Universität von Bristol mit Datenbanksuche, beinhaltet ebenso Seiten für Kinder)
- Palaeos.com (Umfassendes privates Projekt mit hohem wissenschaftlichem Anspruch, über Dinosaurier und die gesamte Lebewelt der Erdgeschichte)
- Dinosauria On-Line (Privates Projekt mit detaillierten Informationen und wissenschaftlichem Anspruch zu Dinosauriern und anderen mesozoischen Reptiliengruppen)
- Thescelosaurus! (Privates Projekt mit Kladogrammen und Beschreibungen mit wissenschaftlichem Anspruch von vielen Dinosauriergattungen)
- DinoData.org (Mehrfach empfohlenes privates Projekt mit vielfältigen Informationen auch zu anderen mesozoischen Landwirbeltieren)
- EnchantedLearning.com (Kommerzielle Internetpräsenz ohne Registrierungszwang mit knappen und sachlichen Informationen, richtet sich an Erwachsene und Kinder)
- The Paleobiology Database – Dinosaur facts and figures (Kuriose Statistiken einer wissenschaftlichen Datenbank)
- Bilder
- Skeletal Drawing (Bilder zur Anatomie der Dinosaurier erstellt von dem Paläontologen Scott Hartman)
Einzelnachweise
- ↑ Wilhelm Gemoll: Griechisch-Deutsches Schul- und Handwörterbuch. München/Wien 1965.
- ↑ Randal Irmis: A reappraisal of the phylogeny of early dinosaurs. In: The Palaeontological Association Newsletter. 65, 2007, S. 92–94 (PDF).
- ↑ G. Olshevsky: An annotated checklist of dinosaur species by continent. In: Mesozoic Meanderings. 3, 2000, S. 1–157.
- ↑ David Lambert und the Diagram Group: The Dinosaur Data Book. Avon Books, New York 1990, ISBN 978-0380758968, S. 288.
- ↑ S. C. Wang, P. Dodson: Estimating the Diversity of Dinosaurs. In: Proceedings of the National Academy of Sciences USA. 103, Nr. 37, 2006, S. 13601–13605 (PMID 16954187).
- ↑ Dale A. Russell: China and the lost worlds of the dinosaurian era. In: Historical Biology. 10, 1995, S. 3–12.
- ↑ Thomas R. Holtz: Dinosaurs. Random House, New York 2007, S. 36.
- ↑ Michael J. Benton: Fossil quality and naming dinosaurs. In: Biology Letters. 4, Nr. 6, 2008, S. 729–732.
- ↑ D. E. Fastovsky, D. B. Weishampel: The Evolution and Exctinction of the Dinosaurs. Cambridge University Press, Cambridge 1996, ISBN 0-521-44496-9.
- ↑ a b c d Michael J. Benton, Dabid B. Weishampel, Peter Dodson: The Dinosauria. 2. Auflage. University of California Press, Berkeley 2004, ISBN 0-520-24209-2, S. 7–19.
- ↑ M. C. Langer, F. Abdala, M. Richter, M. J. Benton: A sauropodomorph dinosaur from the Upper Triassic (Carnian) of southern Brazil. In: Comptes Rendus de l’Academie des Sciences, Paris: Sciences de la terre et des planètes. 329, 1999, S. 511–517.
- ↑ Richard Cowen: History of Life. 4. Auflage. Blackwell Publishing, ISBN 1-405-11756-7, S. 151–175.
- ↑ T. Kubo, M. J. Benton: Evolution of hindlimb posture in archosaurs: limb stresses in extinct vertebrates. In: Palaeontology. 50, Nr. 6, 2007, S. 1519–1529.
- ↑ H. G. Seeley: On the classification of the fossil animals commonly named Dinosauria. In: Proc R Soc London. 43, Royal Society, 1887, S. 165–171.
- ↑ A. S. Romer: Osteology of the Reptiles. University of Chicago, 1956.
- ↑ R. T. Bakker, P. Galton: Dinosaur monophyly and a new class of vertebrates. In: Nature. 248, 1974, S. 168–172, doi:10.1038/248168a0 (http://www.nature.com/nature/journal/v248/n5444/abs/248168a0.html).
- ↑ L. R. Kump, A. Pavlov, M. A. Arthur: Massive release of hydrogen sulfide to the surface ocean and atmosphere during intervals of oceanic anoxia. In: Geology. 33, Nr. 5, 2005, S. 397–400, doi:10.1130/G21295.1.
- ↑ L. H. Tanner, S. G. Lucas und M. G. Chapman: Assessing the record and causes of Late Triassic extinctions. In: Earth-Science Reviews. 65, Nr. 1-2, 2004, S. 103–139, doi:10.1016/S0012-8252(03)00082-5 ([http://nmnaturalhistory.org/pdf_files/TJB.pdf PDF).
- ↑ P. C. Sereno: The evolution of dinosaurs. In: Science. 284, Nr. 5423, 1999, S. 2137–2147, doi:10.1126/science.284.5423.2137.
- ↑ P. C. Sereno, C. A. Forster, R. R. Rogers, A. M. Monetta: Primitive dinosaur skeleton from Argentina and the early evolution of Dinosauria. In: Nature. 361, 1993, S. 64–66, doi:10.1038/361064a0.
- ↑ a b c d Thomas R., Jr. Holtz, Ralph E. Chapman, Matthew C. Lamanna: The Dinosauria. 2. Auflage. University of California Press, Berkeley 2004, ISBN 0-520-24209-2, S. 627–642.
- ↑ a b c d e David E. Fastovsky, Joshua B. Smith, David B. Weishampel, Peter Dodson: The Dinosauria. 2. Auflage. University of California Press, Berkeley 2004, ISBN 0-520-24209-2, S. 614–626.
- ↑ P. C. Sereno, J. A. Wilson, L. M. Witmer, J. A. Whitlock, A. Maga, O. Ide, T. A. Rowe: Structural extremes in a Cretaceous dinosaur. In: PLoS ONE. 2, Nr. 11, 2007, S. e1230, doi:10.1371/journal.pone.0001230 (http://www.plosone.org/article/fetchArticle.action?articleURI=info:doi/10.1371/journal.pone.0001230).
- ↑ V. Prasad, C. A. E. Strömberg, H. Alimohammadian, A. Sahni: Dinosaur coprolites and the early evolution of grasses and grazers. In: Science. 310, Nr. 5751, 2005, S. 1170–1180, doi:10.1126/science.1118806.
- ↑ a b K. Padian; D. B. Weishampel, P. Dodson, H. Osmólska (Hrsg.): The Dinosauria. 2. Auflage. University of California Press, Berkeley 2004, ISBN 0-520-24209-2, S. 210–231.
- ↑ John H. Ostrom: The ancestry of birds. In: Nature. 242, Nr. 5393, 1973, S. 136, doi:10.1038/242136a0.
- ↑ G. Mayr, B. Pohl, D. S. Peters: A Well-Preserved Archaeopteryx Specimen with Theropod Features. In: Science. 310, Nr. 5753, 2005, S. 1483–1486, doi:10.1126/science.1120331.
- ↑ Xu X., M.A. Norell, Kuang X., Wang X., Zhao Q., Jia C.: Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids. In: Nature. 431, Nr. 7009, 2004, S. 680–684 (PMID 15470426).
- ↑ U. B. Göhlich, L. M. Chiappe: A new carnivorous dinosaur from the Late Jurassic Solnhofen archipelago. In: Nature. 440, 2006, S. 329–332.
- ↑ P. Burgers, L. M. Chiappe: The wing of Archaeopteryx as a primary thrust generator. In: Nature. Nr. 399, 1999, S. 60–62 ([http://www.stephenjaygould.org/ctrl/news/file013.html Online).
- ↑ R. Cowen: History of Life. Blackwell Science.
- ↑ a b S. Chatterjee, R. J. Templin: Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui. In: Proceedings of the National Academy of Sciences. 104, Nr. 5, 2007, S. 1576–1580 ([http://www.pnas.org/cgi/reprint/0609975104v1.pdf PDF).
- ↑ J. Peczkis: Implications of body-mass estimates for dinosaurs. In: Journal of Vertebrate Paleontology. Bd. 14, Nr. 4, 1994, S. 520–533.
- ↑ Anatomy and evolution. National Museum of Natural History, abgerufen am 21. November 2007 (englisch).
- ↑ a b Kenneth Carpenter: Biggest of the Big: A critical re-evaluation of the mega-sauropod Amphicoelias fragillimus Cope, 1878. In: Foster, Lucas (Hrsg.): Paleontology and Geology of the Upper Jurassic Morrison Formation. Bulletin 36, New Mexico Museum of Natural History and Science, 2006, S. 131–138 ([https://scientists.dmns.org/sites/kencarpenter/PDFs of publications/Amphicoelias.pdf PDF).
- ↑ John S. McIntosh, M. K. Brett-Surman, James O. Farlow: Sauropods. In: Farlow, Brett-Surman (Hrsg.): The complete Dinosaur. Indiana University Press, 1997, ISBN 0-253-21313-4, S. 281.
- ↑ E. H. Colbert: Men and Dinosaurs: The Search in Field and Laboratory. E. P. Dutton & Company, New York 1968, ISBN 0-14-021288-4, S. vii + 283.
- ↑ David Lovelance, Scott Hartman, William Wahl: Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of Diplodocoid phylogeny. In: Arquivos do Museu Nacional. 65, Nr. 4, Rio de Janeiro 2007, ISSN 0365-4508, S. 527–544.
- ↑ C. dal Sasso, S. Maganuco, E. Buffetaut, M. A. Mendez: New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities. In: Journal of Vertebrate Paleontology. Bd. 25, Nr. 4, 2006, S. 888–896.
- ↑ J. Yans, J. Dejax, D. Pons, C. Dupuis, P. Taquet: Implications paléontologiques et géodynamiques de la datation palynologique des sédiments à faciès wealdien de Bernissart (bassin de Mons, Belgique). In: Comptes Rendus Palevol. 4, Nr. 1-2, 2005, S. 135–150, doi:10.1016/j.crpv.2004.12.003.
- ↑ John Horner, David Weishampel, Catherine Forster: Hadrosauridae. In: Weishampel, Dodson, Osmólska (Hrsg.): The Dinosauria. University of California Press, 2004, ISBN 978-0-520-25408-4, S. 25–46.
- ↑ J. J. Day, P. Upchurch: Sauropod trackways, evolution, and behavior. In: Science. 296, 2002, S. 1659, doi:10.1126/science.107016.
- ↑ Don Lessem, Donald F. Glut: The Dinosaur Society’s Dinosaur Encyclopedia. Random House, 1993, ISBN 0-679-41770-2, S. 19–20.
- ↑ W. D. Maxwell, J. H. Ostrom: Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations. In: Journal of Vertebrate Paleontology. Bd. 15, Nr. 4, 1995, S. 707–712 (Abstract ).
- ↑ Brian T. Roach, Daniel L. Brinkman: A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs. In: Bulletin of the Peabody Museum of Natural History. 48, Nr. 1, 2007, S. 103–138.
- ↑ J. R. Horner, R. Makela: Nest of juveniles provides evidence of family-structure among dinosaurs. In: Nature. Bd. 282, Nr. 5736, 1979, S. 296–298.
- ↑ Oviraptor nesting Nester von Oviraptor oder Protoceratops?
- ↑ Meng Qingjin, Liu Jinyuan, David Varricchio, Timothy Huang, Gao Chunling: Parental care in an ornithischian dinosaur. In: Nature. 431, 2004, S. 145–146.
- ↑ Luis Chiappe, Frankie Jackson, Rodolfo Coria, Lowell Dingus: Nesting Titanosaurs from Auca Mahuevo and Adjacent Sites – Understanding Sauropod Reproductive Behavior and Embryonic Development. In: Rogers, Wilson (Hrsg.): The Sauropods: Evolution and Paleobiology. University of California Press, 2005, ISBN 0-520-24623-3, S. 285–302.
- ↑ Darren H. Tanke, Philip J. Currie: Head-biting behavior in theropod dinosaurs: paleopathological evidence. In: Gaia. 15, 1998, ISSN 0871-5424, S. 167–184 (PDF).
- ↑ James A. Hopson: The evolution of cranial display structures in hadrosaurian dinosaurs. In: Paleobiology. 1, Nr. 1, 1975, S. 21–43.
- ↑ Carl F. Diegert, Thomas E. Williamson: A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen. In: Journal of Vertebrate Paleontology. Bd. 18, Nr. 3, Suppl., 1998, S. 38A.
- ↑ X. Xu, M. A. Norell: A new troodontid dinosaur from China with avian-like sleeping posture. In: Nature. 431, 2004, S. 838–841, doi:10.1038/nature02898.
- ↑ The Fighting Dinosaurs. American Museum of Natural History, abgerufen am 5. Dezember 2007.
- ↑ Kenneth Carpenter: Evidence of predatory behavior by theropod dinosaurs. In: Gaia. 15, 1998, S. 135–144 (Online).
- ↑ Raymond Rogers, David Krause, Kristina Curry Rogers: Cannibalism in the Madagascan dinosaur Majungatholus atopus. In: Nature. 422, Nr. 6931, 2007, S. 515–518, doi:10.1038/nature01532.
- ↑ D. J. Varricchio, A. J. Martin und Y. Katsura: First trace and body fossil evidence of a burrowing, denning dinosaur. In: Proceedings of the Royal Society B: Biological Sciences. 274, Nr. 1616, 2007, S. 1361–1368, doi:10.1098/rspb.2006.0443.
- ↑ F. Zhang, Z. Zhou, X. Xu, X. Wang: A juvenile coelurosaurian theropod from China indicates arboreal habits. In: Naturwissenschaften. 89, Nr. 9, 2002, S. 394–398, doi:10.1007 /s00114-002-0353-8.
- ↑ R. M. Alexander: Dinosaur biomechanics. In: Proceedings of the Royal Society of Biological Sciences. 273, Nr. 1596, 2006, S. 1849–1855, doi:10.1098/rspb.2006.3532.
- ↑ A. Goriely, T. McMillen: Shape of a cracking whip. In: Physical Review Letters. 88, Nr. 24, 2002, S. 244301, doi:10.1103/PhysRevLett.88.244301.
- ↑ D. M. Henderson: Effects of stomach stones on the buoyancy and equilibrium of a floating crocodilian: A computational analysis. In: Canadian Journal of Zoology. Bd. 81, Nr. 8, 2003, S. 1346–1357, doi:10.1139/z03-122.
- ↑ Rubén Ezquerra, Stéfan Doublet, Loic Costeur, Peter M. Galton & Felix Pérez-Lorente: Were non-avian theropod dinosaurs able to swim? Supportive evidence from an Early Cretaceous trackway, Cameros Basin (La Rioja, Spain). In: Geology, 35, 2007; Seiten 507-510
- ↑ T. H. Rich, P. Vickers-Rich, R. A. Gangloff: Polar Dinosaurs. In: Science. 295, 2002, S. 979–980.
- ↑ K. M. Parsons: Drawing Out Leviathan. Indiana University Press, 2001, ISBN 0-253-33937-5, S. 22–48.
- ↑ Robert A. Eagle et al.: Dinosaur Body Temperatures Determined from Isotopic (13C-18O) Ordering in Fossil Biominerals. In: Science, Online-Vorabveröffentlichung vom 23. Juni 2011, doi:10.1126/science.1206196
idw-online.de vom 23. Juni 2011: „Ein Thermometer für Dinosaurier.“ - ↑ P. M. O’Connor, LPAM Claessens: Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs. In: Nature. 436, 2005, S. 253–256, doi:10.1038/nature03716.
- ↑ Mathew Wedel: Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs. In: Paleobiology. 29, Nr. 2, 2003, S. 243–255.
- ↑ P. E. Fisher, D. A. Russell, M. K. Stoskopf, R. E. Barrick, M. Hammer, A. A. Kuzmitz: Cardiovascular evidence for an intermediate or higher metabolic rate in an ornithischian dinosaur. In: Science. 288, Nr. 5465, 2000, S. 503–505, doi:10.1126/science.288.5465.503.
- ↑ W. J. Hillenius, J. A. Ruben: The evolution of endothermy in terrestrial vertebrates: Who? when? why?. In: Physiological and Biochemical Zoology. 77, Nr. 6, 2004, S. 1019–1042, doi:10.1086/425185.
- ↑ T. Rowe, E. F. McBride, P. C. Sereno: Dinosaur with a Heart of Stone. In: Science. 291, Nr. 5505, 2001, S. 783, doi:10.1126/science.291.5505.783a.
- ↑ Chinsamy, Anusuya, Hillenius, J. Willem: Physiology of nonavian dinosaurs. In: The Dinosauria. 2. Auflage. 2004, S. 643–659.
- ↑ M. H. Schweitzer, J. L. Wittmeyer, J. R. Horner: Gender-specific reproductive tissue in ratites and Tyrannosaurus rex. In: Science. 308, 2005, S. 1456–1460, doi:10.1126/science.1112158 (http://www.sciencemag.org/cgi/content/abstract/308/5727/1456).
- ↑ Andrew H. Lee, Sarah Werning: Sexual maturity in growing dinosaurs does not fit reptilian growth models. In: Proceedings of the National Academy of Sciences. 105, Nr. 2, 2008, S. 582–587, doi:10.1073/pnas.0708903105 ([http://www.pnas.org/content/105/2/582.full.pdf+html PDF).
- ↑ C. Dal Sasso, M. Signore: Exceptional soft-tissue preservation in a theropod dinosaur from Italy. In: Nature. 292, Nr. 6674, 1998, S. 383–387, doi:10.1038/32884.
- ↑ M. H. Schweitzer, J. L. Wittmeyer, J. R. Horner: Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex. In: Science. 307, 2005, S. 1952–1955.
- ↑ T. G. Kaye, G. Gaugler, Z. Sawlowicz: Dinosaurian Soft Tissues Interpreted as Bacterial Biofilms. In: PLoS ONE. 3, Nr. 7, e2808, 2008, doi:10.1371/journal.pone.0002808.
- ↑ H. Wang, Z. Yan, D. Jin: Reanalysis of published DNA sequence amplified from Cretaceous dinosaur egg fossil. In: Molecular Biology and Evolution. 14, 1997, S. 589–591 (Online).
- ↑ B. S. Chang, K. Jönsson, M. A. Kazmi, M. J. Donoghue, T. P. Sakmar: Recreating a Functional Ancestral Archosaur Visual Pigment. In: Molecular Biology and Evolution. 19, Nr. 9, 2002, S. 1483–1489 ([1]).
- ↑ G. Embery, A. C. Milner, R. J. Waddington, R. C. Hall, M. S. Langley, A. M. Milan: Identification of proteinaceous material in the bone of the dinosaur Iguanodon. In: Connect Tissue Res. 44, Nr. Suppl 1, 2003, S. 41–6, PMID 12952172.
- ↑ M. H. Schweitzer, M. Marshall, K. Carron, D. S. Bohle, S. C. Busse, E. V. Arnold, D. Barnard, J. R. Horner, J. R. Starkey: Heme compounds in dinosaur trabecular bone. In: Proc Natl Acad Sci U S A.. 94, Nr. 12, 1997, S. 6291–6, PMID 9177210.
- ↑ a b David Archibald, David Fastovsky: Dinosaur extinction. In: Weishampel, Dodson, Osmólska (Hrsg.): The Dinosauria. University of California Press, 2004, ISBN 978-0-520-25408-4.
- ↑ Adriana Ocampo, Vivi Vajda, Eric Buffetaut: Unravelling the Cretaceous–Paleogene (KT) Turnover, Evidence from Flora, Fauna and Geology in Biological Processes Associated with Impact Events. In: Charles Cockell, Iain Gilmour, Christian Koeberl (Hrsg.): Biological Processes Associated with Impact Events. Springer, Berlin, Heidelberg 2006, ISBN 978-3-540-25735-6 (Abstract).
- ↑ J. Fassett, R. A. Zielinski, J. R. Budahn: Dinosaurs that did not die; evidence for Paleocene dinosaurs in the Ojo Alamo Sandstone, San Juan Basin, New Mexico. In: C. Koeberl, K. MacLeod (Hrsg.): Catastrophic events and mass extinctions; impacts and beyond (Geological Society of America – Special Paper). 356, 2002, S. 307–336.
- ↑ Robert Sullivan: No Paleocene dinosaurs in the San Juan Basin, New Mexico. In: Geological Society of America, Abstracts with Programs. 35, Nr. 5, 2003, S. 15.
- ↑ David E. Fastovsky, Yifan Huang, Jason Hsu, Jamie Martin-McNaughton, Peter M. Sheehan, David B. Weishampel: Shape of Mesozoic dinosaur richness. In: Geology. 32, Nr. 10, 2004, S. 877–880, doi:10.1130/G20695.1.
- ↑ G. T. Lloyd, K. E. Davis, D. Pisani, et al.: Dinosaurs and the Cretaceous Terrestrial Revolution. In: Proceedings of the Royal Society: Biology. 2008, doi:10.1098/rspb.2008.0715 ([http://journals.royalsociety.org/content/7k63203q852h4006/fulltext.pdf PDF).
- ↑ Zhiming Dong: Dinosaurian Faunas of China. China Ocean Press, Beijing 1992, ISBN 3-540-52084-8.
- ↑ Dinosaur bones 'used as medicine'. BBC News, 2007, abgerufen am 6. Juni 2007.
- ↑ Hans-Dieter Sues; James Orville Farlow, M. K. Brett-Surman (Hrsg.): The Complete Dinosaur. Indiana University Press, Bloomington 1997, ISBN 0-253-33349-0, European Dinosaur Hunters, S. 14.
- ↑ Deborah Cadbury: Dinosaurier Jäger. Rowohlt Verlag, 2000, ISBN 3-498-00924-9.
- ↑ T. Holmes: Fossil Feud: The Bone Wars of Cope and Marsh, Pioneers in Dinosaur Science. Silver Burdett Press, 1996, ISBN 978-0382391477.
- ↑ David Norman: Dinosaurier. Bertelsmann Verlag, München 1991 (übersetzt von Christoph Arndt), ISBN 3-570-02250-1.
- ↑ Kenneth Carpenter: Dinosaurs as Museum Exhibits. In: James Farlow, Brett-Surman (Hrsg.): The complete dinosaur. Indiana University Press, 1997, ISBN 0-253-21313-4.
- ↑ Smithsonian National Museum of Natural History – Department of Paleobiology
- ↑ A. Geiger: Die Faszination für Kulturen vor der Kultur: Ägyptomanie und Dinosaurierkult – 2. Was macht Dinosaurier interessant?. In: Stuttgarter Unikurier. Nr. 80, November 1998 (Online).


Dieser Artikel wurde in die Liste der lesenswerten Artikel aufgenommen. Kategorien:- Wikipedia:Lesenswert
- Dinosaurier


















Wikimedia Foundation.